Studies Relating to the Theory of Panspermia
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Modelling Panspermia in the TRAPPIST-1 System
October 13, 2017 Modelling panspermia in the TRAPPIST-1 system James A. Blake1,2*, David J. Armstrong1,2, Dimitri Veras1,2 Abstract The recent ground-breaking discovery of seven temperate planets within the TRAPPIST-1 system has been hailed as a milestone in the development of exoplanetary science. Centred on an ultra-cool dwarf star, the planets all orbit within a sixth of the distance from Mercury to the Sun. This remarkably compact nature makes the system an ideal testbed for the modelling of rapid lithopanspermia, the idea that micro-organisms can be distributed throughout the Universe via fragments of rock ejected during a meteoric impact event. We perform N-body simulations to investigate the timescale and success-rate of lithopanspermia within TRAPPIST-1. In each simulation, test particles are ejected from one of the three planets thought to lie within the so-called ‘habitable zone’ of the star into a range of allowed orbits, constrained by the ejection velocity and coplanarity of the case in question. The irradiance received by the test particles is tracked throughout the simulation, allowing the overall radiant exposure to be calculated for each one at the close of its journey. A simultaneous in-depth review of space microbiological literature has enabled inferences to be made regarding the potential survivability of lithopanspermia in compact exoplanetary systems. 1Department of Physics, University of Warwick, Coventry, CV4 7AL 2Centre for Exoplanets and Habitability, University of Warwick, Coventry, CV4 7AL *Corresponding author: [email protected] Contents Universe, and can propagate from one location to another. This interpretation owes itself predominantly to the works of William 1 Introduction1 Thompson (Lord Kelvin) and Hermann von Helmholtz in the 1.1 Mechanisms for panspermia...............2 latter half of the 19th Century. -

Prebiological Evolution and the Metabolic Origins of Life
Prebiological Evolution and the Andrew J. Pratt* Metabolic Origins of Life University of Canterbury Keywords Abiogenesis, origin of life, metabolism, hydrothermal, iron Abstract The chemoton model of cells posits three subsystems: metabolism, compartmentalization, and information. A specific model for the prebiological evolution of a reproducing system with rudimentary versions of these three interdependent subsystems is presented. This is based on the initial emergence and reproduction of autocatalytic networks in hydrothermal microcompartments containing iron sulfide. The driving force for life was catalysis of the dissipation of the intrinsic redox gradient of the planet. The codependence of life on iron and phosphate provides chemical constraints on the ordering of prebiological evolution. The initial protometabolism was based on positive feedback loops associated with in situ carbon fixation in which the initial protometabolites modified the catalytic capacity and mobility of metal-based catalysts, especially iron-sulfur centers. A number of selection mechanisms, including catalytic efficiency and specificity, hydrolytic stability, and selective solubilization, are proposed as key determinants for autocatalytic reproduction exploited in protometabolic evolution. This evolutionary process led from autocatalytic networks within preexisting compartments to discrete, reproducing, mobile vesicular protocells with the capacity to use soluble sugar phosphates and hence the opportunity to develop nucleic acids. Fidelity of information transfer in the reproduction of these increasingly complex autocatalytic networks is a key selection pressure in prebiological evolution that eventually leads to the selection of nucleic acids as a digital information subsystem and hence the emergence of fully functional chemotons capable of Darwinian evolution. 1 Introduction: Chemoton Subsystems and Evolutionary Pathways Living cells are autocatalytic entities that harness redox energy via the selective catalysis of biochemical transformations. -

Why Can't Cosmology Be More Open?
ISSN: 2641-886X International Journal of Cosmology, Astronomy and Astrophysics Opinion Article Open Access Why can’t Cosmology be more open? Jayant Narlikar* Inter-University Centre for Astronomy and Astrophysics, Ganeshkhind, Post Bag 4, Pune-411007, India Article Info Keywords: Cosmology, Galaxy, Big Bang, Spectrum *Corresponding author: More than two millennia back, Pythagoreans believed that the Earth went round a Jayant Narlikar central fire, with the Sun lying well outside its orbit. When sceptics asked,” Why can’t we Emeritus Professor see the fire?”, theorists had to postulate that there was a ‘counter-Earth’ going around Inter-University Centre for Astronomy and Astrophysics the central fire in an inner orbit that blocked our view of the fire. The sceptics asked Ganeshkhind, Post Bag 4 again: why don’t we see this ‘counter-Earth’? The theorists replied that this happened Pune, India because Greece was facing away from it. In due course this explanation too was also Tel: +91-20-25604100 shot down by people sailing around and looking from other directions. Fax: +91-20-25604698 E-mail: [email protected] I have elaborated this ancient episode because it holds a moral for scientists. When you are on the wrong track, you may have to invoke additional assumptions, like the Received: November 5, 2018 counter-Earth, to prop up your original theory against an observed fact. If there is no Accepted: November 15, 2018 other independent support for these assumptions, the entire structure becomes suspect. Published: January 2, 2019 The scientific approach then requires a critical re-examination of the basic paradigm. -

6 Dynamical Generalizations of the Drake Equation: the Linear and Non-Linear Theories
6 Dynamical Generalizations of the Drake Equation: The Linear and Non-linear Theories Alexander D. Panov Abstract The Drake equation pertains to the essentially equilibrium situation in a popu- lation of communicative civilizations of the Galaxy, but it does not describe dynamical processes which can occur in it. Both linear and non-linear dynam- ical population analyses are built out and discussed instead of the Drake equa- tion. Keywords: SETI, the Drake equation, linear population analysis, non-linear population analysis. Introduction Communicative civilizations (CCs) are the ones which tend to send messages to other civilizations and are able to receive and analyze messages from other civili- zations. The crucial question of the SETI problem is how far the nearest CC from us is. Its answer depends on the number of CCs existing in the Galaxy at present. Fig. 1 shows how the distance between the Sun and the nearest CC depends on the number of CCs in the Galaxy. The calculation was made by us by the Monte Carlo method with the use of a realistic model of the distribution of stars in the Galaxy (Allen 1973) and the actual location of the Sun in the Galaxy (8.5 kpc from the center of the Galaxy). The best known way to answer the question about the number of CCs is the formula by F. Drake N R f n f f f L C * p e l i c , (Eq. 1) where R∗ is a star-formation rate in the Galaxy averaged with respect to all time of its existence, fp is the part of stars with planet systems, nе is the average number of planets in systems suitable for life, fl is the part of planet on which life did appear, fi is the part of planets on which intelligent forms of life devel- oped, fc is the part of planets on which life reached the communicative phase, L is the average duration of the communicative phase. -
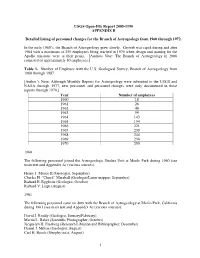
USGS Open-File Report 2005-1190, Appendix A
USGS Open-File Report 2005-1190 APPENDIX B Detailed listing of personnel changes for the Branch of Astrogeology from 1960 through 1972. In the early 1960’s, the Branch of Astrogeology grew slowly. Growth was rapid during and after 1964 with a maximum of 250 employees being reached in 1970 when design and training for the Apollo missions were at their peaks. [Authors Note: The Branch of Astrogeology in 2006 consisted of approximately 80 employees.] Table 1. Number of Employee with the U.S. Geological Survey, Branch of Astrogeology from 1960 through 1987 [Author’s Note: Although Monthly Reports for Astrogeology were submitted to the USGS and NASA through 1977, new personnel and personnel changes were only documented in these reports through 1970.] Year Number of employees 1960 18 1961 26 1962 40 1963 59 1964 143 1965 154 1966 221 1967 239 1968 244 1969 234 1970 250 1960 The following personnel joined the Astrogeologic Studies Unit at Menlo Park during 1960 (see main text and Appendix A) (various sources): Henry J. Moore II (Geologist, September) Charles H. “Chuck” Marshall (Geologist/Lunar mapper; September) Richard E. Eggleton (Geologist, October) Richard V. Lugn (August) 1961 The following personnel came on duty with the Branch of Astrogeology at Menlo Park, California during 1961 (see main text and Appendix A) (various sources): David J. Roddy (Geologist; January/February) Martin L. Baker (Scientific Photographer; October) Jacquelyn H. Freeberg (Research Librarian and Bibliographer; December) Daniel J. Milton (Geologist; August) Carl H. Roach (Geophysicist; August) 1 Maxine Burgess (Secretary) 1962 The following was taken from the Branch of Astrogeology Monthly Report for April 1962 from Chief, Branch of Astrogeology to V.E. -

Messengers from Outer Space
The complete absence of national laboratories for standardizing radiation measurements and the lack of physics departments in most radiotherapy centres in Latin America warrant the setting up of one or more regional dosimetry facilities, the functions of which would be primarily to calibrate dosimeters; to provide local technical assistance by means of trained staff; to check radiation equipment and dosimeters; to arrange a dose-intercomparison service; and to co-operate with local personnel dosimetry services. To be most effective, this activity should be in the charge of local personnel, with some initial assistance in the form of equipment and experts provided by the Agency. Although the recommendations were presented to the IAEA as the or ganization responsible for the setting up of the panel, the participants re commended that the co-operation of the World Health Organization and the Pan-American Health Organization should be invited. It was also suggested that the panel report be circulated to public health authorities in the countries represented. MESSENGERS FROM OUTER SPACE Although no evidence has yet been confirmed of living beings reaching the earth from space, it has been estimated that hundreds of tons of solid matter arrive every day in the form of meteorites or cosmic dust particles. Much is destroyed by heat in the atmosphere but fragments recovered can give valuable information about what has been happening in the universe for billions of years. Some of the results of worldwide research on meteorites were given at a symposium in Vienna during August. During six days of discussions a total of 73 scientific papers was present ed and a special meeting was held for the purpose of improving international co-operation in the research. -

Cometary Panspermia a Radical Theory of Life’S Cosmic Origin and Evolution …And Over 450 Articles, ~ 60 in Nature
35 books: Cosmic origins of life 1976-2020 Physical Sciences︱ Chandra Wickramasinghe Cometary panspermia A radical theory of life’s cosmic origin and evolution …And over 450 articles, ~ 60 in Nature he combined efforts of generations supporting panspermia continues to Prof Wickramasinghe argues that the seeds of all life (bacteria and viruses) Panspermia has been around may have arrived on Earth from space, and may indeed still be raining down some 100 years since the term of experts in multiple fields, accumulate (Wickramasinghe et al., 2018, to affect life on Earth today, a concept known as cometary panspermia. ‘primordial soup’, referring to Tincluding evolutionary biology, 2019; Steele et al., 2018). the primitive ocean of organic paleontology and geology, have painted material not-yet-assembled a fairly good, if far-from-complete, picture COMETARY PANSPERMIA – cultural conceptions of life dating back galactic wanderers are normal features have argued that these could not into living organisms, was first of how the first life on Earth progressed A SOLUTION? to the ideas of Aristotle, and that this of the cosmos. Comets are known to have been lofted from the Earth to a coined. The question of how from simple organisms to what we can The word ‘panspermia’ comes from the may be the source of some of the have significant water content as well height of 400km by any known process. life’s molecular building blocks see today. However, there is a crucial ancient Greek roots ‘sperma’ meaning more hostile resistance the idea of as organics, and their cores, kept warm Bacteria have also been found high in spontaneously assembled gap in mainstream understanding - seed, and ‘pan’, meaning all. -
The Water on Mars Vanished. This Might Be Where It Went
The water on Mars vanished. This might be where it went. timesofindia.indiatimes.com/home/science/the-water-on-mars-vanished-this-might-be-where- it-went-/articleshow/81599751.cms NYT News Service | Mar 20, 2021, 10:32 IST Mars was once wet, with an ocean’s worth of water on its surface. Today, most of Mars is as dry as a desert except for ice deposits in its polar regions. Where did the rest of the water go? Some of it disappeared into space. Water molecules, pummeled by particles of solar wind, broke apart into hydrogen and oxygen atoms, and those, especially the lighter hydrogen atoms, sped out of the atmosphere, lost to outer space. A tall outcropping of rock, with layered deposits of sediments in the distance, marking a remnant of an ancient, long-vanished river delta in Jezero Crater, are pictured in this undated image taken by NASA's Mars rover Perseverance. (Reuters) But most of the water, a new study concludes, went down, sucked into the red planet’s rocks. And there it remains, trapped within minerals and salts. Indeed, as much as 99% of the water that once flowed on Mars could still be there, the researchers estimated in a paper published this week in the journal Science. Data from the past two decades of robotic missions to Mars, including NASA ’s Curiosity rover and the Mars Reconnaissance Orbiter, showed a wide distribution of what geologists call hydrated minerals. “It became very, very clear that it was common and not rare to find evidence of water alteration,” said Bethany Ehlmann, a professor of planetary science at the California Institute of Technology and one of the authors of the paper. -

The Protection of Frequencies for Radio Astronomy 1
JOURNAL OF RESEARCH of the National Bureau of Standards-D. Radio Propagation Vol. 67D, No. 2, March- April 1963 b The Protection of Frequencies for Radio Astronomy 1 R. 1. Smith-Rose President, International Scientific Radio Union (R eceived November 5, 1962) The International T elecommunications Union in its Geneva, 1959 R adio R egulations r recognises the Radio Astronomy Service in t he two following definitions: N o. 74 Radio A st1" onomy: Astronomy based on t he reception of waves of cos mi c origin. No. 75 R adio A st1"onomy Se1"vice: A service involving the use of radio astronomy. This service differs, however, from other r adio services in two important respects. 1. It does not itself originate any radio waves, and therefore causes no interference to any other service. L 2. A large proportion of its activity is conducted by the use of reception techniques which are several orders of magnit ude )]).ore sensitive than those used in other ra dio services. In order to pursue his scientific r esearch successfully, t he radio astronomer seeks pro tection from interference first, in a number of bands of frequencies distributed t hroughout I t he s p ~ct run:; and secondly:. 1~10r e complete and s p ec i~c prote.ction fOl: t he exact frequency bands III whIch natural radIatIOn from, or absorptIOn lD, cosmIc gases IS known or expected to occur. The International R egulations referred to above give an exclusive all ocation to one freq uency band only- the emission line of h ydrogen at 1400 to 1427 Mc/s. -

Cosmochemistry Cosmic Background Radia�On
6/10/13 Cosmochemistry Cosmic background radiaon Dust Anja C. Andersen Niels Bohr Instute University of Copenhagen hp://www.dark-cosmology.dk/~anja Hauser & Dwek 2001 Molecule formation on dust grains Multiwavelenght MW 1 6/10/13 Gas-phase element depleons in the Concept of dust depleon interstellar medium The depleon of an element X in the ISM is defined in terms of (a logarithm of) its reducon factor below the expected abundance relave to that of hydrogen if all of the atoms were in the gas phase, [Xgas/H] = log{N(X)/N(H)} − log(X/H) which is based on the assumpon that solar abundances (X/H)are good reference values that truly reflect the underlying total abundances. In this formula, N(X) is the column density of element X and N(H) represents the column density of hydrogen in both atomic and molecular form, i.e., N(HI) + 2N(H2). The missing atoms of element X are presumed to be locked up in solids within dust grains or large molecules that are difficult to idenfy spectroscopically, with fraconal amounts (again relave to H) given by [Xgas/H] (Xdust/H) = (X/H)(1 − 10 ). Jenkins 2009 Jenkins 2009 2 6/10/13 Jenkins 2009 Jenkins 2009 The Galacc Exncon Curve Extinction curves measure the difference in emitted and observed light. Traditionally measured by comparing two stars of the same spectral type. Galactic Extinction - empirically determined: -1 -1 <A(λ)/A(V)> = a(λ ) + b(λ )/RV (Cardelli et al. 1999) • Bump at 2175 Å (4.6 µm-1) • RV : Ratio of total to selective extinction in the V band • Mean value is RV = 3.1 (blue) • Low value: RV = 1.8 (green) (Udalski 2003) • High value: RV = 5.6-5.8 (red) (Cardelli et al. -

Exploring Biogenic Dispersion Inside Star Clusters with System Dynamics Modeling
Preprints (www.preprints.org) | NOT PEER-REVIEWED | Posted: 12 October 2020 doi:10.20944/preprints202010.0232.v1 Exploring Biogenic Dispersion Inside Star Clusters with System Dynamics Modeling Authors Javier D. Burgos. Corporación para la Investigacion e Innovación -CIINAS. Contact: [email protected] Diana C. Sierra. Corporación para la Investigacion e Innovación -CIINAS; Bogotá, Colombia. Contact: [email protected] Abstract The discovery of a growing number of exoplanets and even extrasolar systems supports the scientific consensus that it is possible to find other signs of life in the universe. The present work proposes for the first time, an explicit mechanism inspired by the dynamics of biological dispersion, widely used in ecology and epidemiology, to study the dispersion of biogenic units, interpreted as complex organic molecules, between rocky or water exoplanets (habitats) located inside star clusters. The 3 results of the dynamic simulation suggest that for clusters with populations lower than 4 M/ly it is not possible to obtain biogenic worlds after 5 Gyr. Above this population size, biogenic dispersion seems to follow a power law, the larger the density of worlds lesser will be the impact rate (훽.) value to obtain at least one viable biogenic Carrier habitat after 5 Gyr. Finally, when we investigate scenarios by varying β, a well-defined set of density intervals can be defined in accordance to its characteristic β value, suggesting that biogenic dispersion has a behavior of “minimal infective dose” of “minimal biogenic effective” events by interval i.e. once this dose has been achieved, doesn’t matter if additional biogenic impact events occur on the habitat. -

Transcript-Life-Beyond-Earth-Mind-Man
Video Transcript for Archival Research Catalog (ARC) Identifier 649452 Life Beyond Earth and the Mind of Man – 1975 Ashley Montagu: When we speak of life beyond Earth, what we generally mean is of course intelligent life, something resembling our noble selves. It is highly probable that there are such intelligent forms of life in other galaxies in the universe. And it is even more probable that many of these forms are vastly more intelligent than we. Philip Morrison: It is conceivable that a spherical ship will land in front of the Washington Monument and a figure with four antennas and otherwise looking like a professional football player will walk out and demand to see our leader. But I hope very much that the universe of circumstance is wider than the rather shoddy imaginations of science fiction writers for 30 or 40 years. And I’m pretty convinced it is. We’ve not found their guidance so great in any but the most modest activities, like going to the Moon, which is not very much. George Wald: May I reply to that one. Narrator: Life Beyond Earth – George Wald: You see, this is a beautiful exposition of the myths we live with. Narrator: – and the Mind of Man, a symposium on extraterrestrial life and some of its implications for us all. Sponsored jointly by NASA and Boston University and chaired by Professor Richard Berendzen. Richard Berendzen: Let me briefly introduce the panel. On the far side is Dr. Ashley Montagu who’s a world-renowned anthropologist and social biologist. Next to him is Dr.