SARA Project
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Rodeo Program Canadian Senior Pro Rodeo COCHRANE-1SAT Cochrane 05/18/2019
Rodeo Program Canadian Senior Pro Rodeo COCHRANE-1SAT Cochrane 05/18/2019 BARREL RACING 40-49 CALF ROPING 50-59 Contestant City Back # Contestant City Back # 1 APRIL HEEG ACADIA VALLEY, AB 0 1 DUANE GANSKE WETASKIWIN, AB 0 2 TASHA TURNER CALGARY, AB 0 2 MANERD BIRD NANTON, AB 0 3 JULIE BLAND CALGARY, AB 0 3 HAROLD WRIGHT LUNDBRECK, AB 0 4 DONALEE FERGUSON MUNSONAB, AB 0 4 DEAN PEROZAK LETHBRIDGE 0 5 RITA ARTEMENKO WESTLOCK, AB 0 5 CRAIG FITZPATRICK OKOTOKS, AB 0 6 JENNIFER PETERS SUNDRE, AB 0 6 DARYL SUTCLIFF NANTON, AB 0 7 NIKI MAKOFKA PONOKA, AB 0 7 LYLE KATHREIN CLARESHOLM, AB 0 8 NIKKI HOLMES CALGARY, AB 0 8 KELLY CREASY MEDICINE HAT, AB 0 9 ANGIE THOMSON BIG VALLEY, AB 0 9 LYNN TURCATO TABER, AB 0 10 HEATHER PARSONS LOCAL ENTRY, 0 10 VAUGHN STEWART DEWINTON, AB 0 11 KELLY MOYNIHAN BANFF, AB 0 CALF ROPING 60+ BARREL RACING 50-59 Contestant City Back # Contestant City Back # 1 NEIL HOWARD OKOTOKS, AB 0 1 MADELAINE KIMMERLY NANTON, 0 2 LONNY OLSON PONOKA, AB 0 2 TRISH BROWN CALGARY, AB 0 3 KENT MOSHER AUGUSTA, MT 0 3 NADENE ADIE ROCKY VIEW 0 4 DOUG SHIPTON REDMOND, OR 0 4 VAL NELSON COCHRANE, AB 0 5 LORNE LAUSEN STRATHMORE, AB 0 5 RHONDA HENRY GLEICHEN, AB 0 6 GUY PEROZAK CLARESHOLM, AB 0 6 THERESA OLDFORD WINFIELD, AB 0 7 GLEN ADIE ROCKY VIEW 0 7 ANNETTE MCCAUGHAN BRANT, AB 0 8 ROGER GUNSCH THORSBY, AB 0 8 CAROLINE HEFFERNAN CARSELAND, AB 0 9 EVERETT MORTON DEL BONITA, AB 0 9 TAMMY SCHUURMAN DIAMOND CITY, AB 0 10 NEIL WATT CLARESHOLM, AB 0 10 ROSE PEROZAK CLARESHOLM, AB 0 CALF ROPING 68+ 11 CONNIE LEMOINE AIRDRIE, AB 0 Contestant City -

BE IDENTIFIABLE 1 Circlededicated to Raising Quality Charolais CEE Cattle Since
Membership2018-2020 Directory BE IDENTIFIABLE 1 CIRCLEDedicated to Raising Quality Charolais CEE Cattle Since... www.circlecee.com 1971 Bulls Available By Private Treaty Off the Farm! CIRCLE CEE CHAROLAIS The Program You Can Depend On! THE CHOLAK FAMILY | LAMONT, ALBERTA | SSCHOLAK@S HAW.CA Stephen & Deb Cholak - 780-485-7865 | Frank & Elly Cholak - 780-796-2108 2 Welcome Charolais Enthusiasts! Dedicated to Raising Quality Charolais Cattle Since... On behalf of the Alberta Charolais Association Board of Directors and over 210 active CIRCLE CEE Charolais Breeders in the beautiful provinces of British Columbia and Alberta, I would like to welcome you to browse the pages of our Breeders Directory. Thanks to the support of these breeders we have a current snapshot of the operations taking our breed and the entire cattle www.circlecee.com industry into the future. We hope that the information is presented in a readable, easily 1971 accessible format that you will find useful in your next search for Charolais genetics. The Charolais breed has a proven, consistent track record of performance in the feedlot which is only gaining momentum. Charolais influenced calves are simply making cow-calf producers and feedlot owners more money. We are truly the “Identifiable” breed and believe our influence will continue as long as we continue to stay focused on the long term goal of the beef industry – profitability. The Alberta Charolais Association hosts a few key events during the year. In late June we annually host a Breeder’s tour where breeders are showcased in various regions of the two provinces. This complimentary tour has attracted over 200 enthusiasts the past 3 years and is a no miss event. -

County of Stettler No. 06
AAAF SPRING FORUM 2012 AGRICULTURAL FIELDMAN’S DIRECTORY –CURRENT TO April 25, 2012 SOUTH REGION M.D. of Acadia Rick Niwa (AF) Office: (403) 972-3808 Box 30, Acadia Valley Shop: (403) 972-3755 T0J 0A0 Fax: (403) 972-3833 Cell: (403) 664-7114 email [email protected] Cardston County Rod Foggin (AF) Ph: (403) 653-4977 Box 580, Cardston Stephen Bevans (AAF) Fax: (403) 653-1126 T0K 0K0 Cell: (403) 382-8236 (Rod) (403) 634-9474 email: [email protected] [email protected] Municipality of Crowsnest Pass Kim Lutz (AF) Ph: (403)-563-8658 Mail: email: [email protected] Box 600 Crowsnest Pass, AB T0K 0E0 Office: Room 1, MDM Community Center 2802 - 222 Street Bellevue, AB Cypress County Jason Storch (AF) Director Ph: (403) 526-2888 816 2nd Ave, Dunmore Christina Barrieau (AAF) Fax: (403) 526-8958 T1B 0K3 email : [email protected] [email protected] M.D. of Foothills Ron Stead (AF) Ph: (403) 603-5410 (Ron) Box 5605, High River Bree Webb (AAF) Shop: (403) 652-2423 (Bree) T1V 1M7 ext 5446 Fax : (403) 603-5414 email : [email protected] [email protected] County of Forty Mile Dave Matz (AF) Phone (403) 867-3530 Box 160, Foremost Vacant (AAF) fax (403) 867-2242 T0K 0X0 Kevin Jesske (Fieldman’s Asst.) cellular (403) 647-8080 (Dave) email [email protected] [email protected] Lethbridge County Don Bodnar (AF) Ph: (403) 328-5525 905-4th Ave. South Gary Secrist (AAF) shop: (403) 732-5333 Lethbridge T1J 4E4 Terry Mrozowich Fax: (403) 732-4328 Cell : (403) 634-0713 (Don) (403) 634-0680 (Gary) email : [email protected] [email protected] County of Newell Todd Green (AF) Office: (403) 362-2772 Box 130, Brooks Holly White (AAF/Rural Cons. -

Evaluating the Monophyly and Biogeography of Cryptantha (Boraginaceae)
Systematic Botany (2018), 43(1): pp. 53–76 © Copyright 2018 by the American Society of Plant Taxonomists DOI 10.1600/036364418X696978 Date of publication April 18, 2018 Evaluating the Monophyly and Biogeography of Cryptantha (Boraginaceae) Makenzie E. Mabry1,2 and Michael G. Simpson1 1Department of Biology, San Diego State University, San Diego, California 92182, U. S. A. 2Current address: Division of Biological Sciences and Bond Life Sciences Center, University of Missouri, Columbia, Missouri 65211, U. S. A. Authors for correspondence ([email protected]; [email protected]) Abstract—Cryptantha, an herbaceous plant genus of the Boraginaceae, subtribe Amsinckiinae, has an American amphitropical disjunct distri- bution, found in western North America and western South America, but not in the intervening tropics. In a previous study, Cryptantha was found to be polyphyletic and was split into five genera, including a weakly supported, potentially non-monophyletic Cryptantha s. s. In this and subsequent studies of the Amsinckiinae, interrelationships within Cryptantha were generally not strongly supported and sample size was generally low. Here we analyze a greatly increased sampling of Cryptantha taxa using high-throughput, genome skimming data, in which we obtained the complete ribosomal cistron, the nearly complete chloroplast genome, and twenty-three mitochondrial genes. Our analyses have allowed for inference of clades within this complex with strong support. The occurrence of a non-monophyletic Cryptantha is confirmed, with three major clades obtained, termed here the Johnstonella/Albidae clade, the Maritimae clade, and a large Cryptantha core clade, each strongly supported as monophyletic. From these phylogenomic analyses, we assess the classification, character evolution, and phylogeographic history that elucidates the current amphitropical distribution of the group. -

COSEWIC Assessment and Status Report on the Tiny Cryptantha Cryptantha Minima in Canada
COSEWIC Assessment and Status Report on the Tiny Cryptantha Cryptantha minima in Canada THREATENED 2012 COSEWIC status reports are working documents used in assigning the status of wildlife species suspected of being at risk. This report may be cited as follows: COSEWIC. 2012. COSEWIC assessment and status report on the Tiny Cryptantha Cryptantha minima in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. x + 37 pp. (www.registrelep-sararegistry.gc.ca/default_e.cfm). Previous report(s): COSEWIC. 2000. COSEWIC assessment and status report on the tiny cryptanthe Cryptantha minima in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. vi + 18 pp. Smith, B. 1998. COSEWIC status report on the tiny cryptanthe Cryptantha minima in Canada, in COSEWIC assessment and status report on the tiny cryptanthe Cryptantha minima in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 1-18 pp. Production note: COSEWIC would like to acknowledge Sue Michalsky for writing the status report on the Tiny Cryptantha Cryptantha minima in Canada, prepared under contract with Environment Canada. This report was overseen and edited by Bruce Bennett and Erich Haber, Co-chairs of the COSEWIC Vascular Plants Specialist Subcommittee. For additional copies contact: COSEWIC Secretariat c/o Canadian Wildlife Service Environment Canada Ottawa, ON K1A 0H3 Tel.: 819-953-3215 Fax: 819-994-3684 E-mail: COSEWIC/[email protected] http://www.cosewic.gc.ca Également disponible en français sous le titre Ếvaluation et Rapport de situation du COSEPAC sur la Cryptanthe minuscule (Cryptantha minima) au Canada. Cover illustration/photo: Tiny Cryptantha — Source: Environment Canada 2010. -
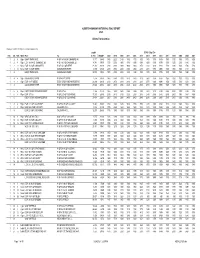
2020 Alberta Highways Historical ESAL Report
ALBERTA HIGHWAY HISTORICAL ESAL REPORT 2020 Alberta Transportation Produced: 24-Feb-2021 By CornerStone Solutions Inc. Length ESAL / Day / Dir Hwy CS TCS Muni From To in Km WAADT 2020 2019 2018 2017 2016 2015 2014 2013 2012 2011 2010 2009 2008 2007 1 2 4 Bigh BANFF PARK GATE W OF 1A NW OF CANMORE WJ 3.777 19480 1830 2220 2140 1760 1710 1600 1470 1370 1610 1550 1550 1530 1370 1400 1 2 8 Bigh E OF 1A NW OF CANMORE WJ W OF 1A S OF CANMORE EJ 4.741 18030 1730 2050 1980 1730 1690 1590 1620 1530 1570 1500 1250 1230 1140 1160 1 2 12 KanC E OF 1A S OF CANMORE EJ W OF 1X S OF SEEBE 23.165 20220 1890 2050 1980 1680 1660 1570 1510 1420 1810 1730 1680 1660 1700 1710 1 2 16 KanC E OF 1X S OF SEEBE KANANASKIS RIVER 0.896 20820 2510 2750 2650 2360 2350 2240 2080 1970 2050 1960 1960 1920 2110 2110 1 2 BANFF PARK GATE KANANASKIS RIVER 32.579 19832 1870 2090 2010 1720 1690 1590 1530 1440 1750 1670 1600 1580 1590 1600 1 4 4 Bigh KANANASKIS RIVER W OF 40 AT SEEBE 3.228 20820 1960 2140 2070 1820 1810 1720 1600 1510 1630 1560 1560 1530 1520 1530 1 4 8 Bigh E OF 40 AT SEEBE E BDY STONY INDIAN RESERVE 22.296 22610 2310 2470 2380 2310 2310 2230 2070 1960 1980 1650 1650 1580 1270 1230 1 4 KANANASKIS RIVER E BDY STONY INDIAN RESERVE 25.524 22384 2260 2430 2340 2240 2240 2160 2010 1900 1940 1640 1630 1570 1300 1270 1 6 4 Rkyv E BDY STONY INDIAN RESERVE W OF JCT 68 3.166 21830 1860 1990 1920 2040 2040 1950 1810 1710 2140 2040 2030 1970 2250 2250 1 6 8 Rkyv E OF JCT 68 W OF 22 S OF COCHRANE 17.235 22340 2330 2410 2330 2520 2530 2640 2490 2360 2410 2010 2000 1960 1880 1800 1 6 E BDY STONY INDIAN RESERVE W OF 22 S OF COCHRANE 20.401 22261 2250 2360 2280 2450 2450 2540 2390 2260 2360 2010 2000 1950 1930 1880 1 8 4 Rkyv E OF 22 S OF COCHRANE W OF 563 W OF CALGARY 11.441 26100 1730 1960 1630 1610 1570 1550 1380 1300 1160 1110 1100 1060 1020 1010 1 8 8 Rkyv E OF 563 W OF CALGARY CALGARY W.C.L. -

Fish Stocking Report, 2020 (Final)
Fish Stocking Report 2020 (Final) Fish stocking managed by the Government of Alberta and the Alberta Conservation Association Updated February 18, 2021 Notes There are no cutthroat trout stocked in the 2020 stocking season, as we will not be operating the Job Lake spawn camp due to COVID-19 restrictions. Average Length = adult fish stocked. Reference Species Stocked Strains Stocked Ploidy Stocked ARGR = Arctic Grayling BEBE = Beity x Beity TLTLJ = Trout Lodge / Jumpers 2N = diploid BKTR = Brook Trout BRBE = Bow River x Beity TLTLK = Trout Lodge / Kamloops 3N = triploid BNTR = Brown Trout CLCL = Campbell Lake TLTLS = Trout Lodge / Silvers AF2N = all female diploid CTTR = Cutthroat Trout JLJL = Job Lake AF3N = all female triploid RNTR = Rainbow Trout LYLY = Lyndon TGTR = Tiger Trout PLPL = Pit Lakes For further information on Fish Stocking visit: https://mywildalberta.ca/fishing/fish-stocking/default.aspx ©2021 Government of Alberta | Published: February 2021 Page 1 of 24 Waterbody Waterbody ATS Species Strain Genotype Average Number Stocking Official Name Common Name Length Stocked Date (2020) ALFORD LAKE SW4-36-8-W5 RNTR Campbell Lake 3N 18 3000 18-May-20 BEAR POND NW36-14-4-W5 RNTR Trout Lodge/Jumpers AF3N 19.7 750 22-Jun-20 BEAUVAIS LAKE SW29-5-1-W5 RNTR Trout Lodge/Jumpers AF3N 16.3 23000 11-May-20 BEAVER LAKE NE16-35-6-W5 RNTR Trout Lodge/Jumpers AF3N 21.3 2500 21-May-20 BEAVER LAKE NE16-35-6-W5 TGTR Beitty/Bow River 3N 16.9 500 02-Sep-20 BEAVER LAKE NE16-35-6-W5 TGTR Beitty/Bow River 3N 20 500 02-Sep-20 BEAVER MINES LAKE NE11-5-3-W5 -

Evolution of Flowering Time in the Tetraploid Capsella Bursa-Pastoris (Brassicaceae)
Digital Comprehensive Summaries of Uppsala Dissertations from the Faculty of Science and Technology 367 Evolution of Flowering Time in the Tetraploid Capsella bursa-pastoris (Brassicaceae) TANJA SLOTTE ACTA UNIVERSITATIS UPSALIENSIS ISSN 1651-6214 UPPSALA ISBN 978-91-554-7024-1 2007 urn:nbn:se:uu:diva-8311 ! " #$$" $%$$ & & & ' ( ) * ( + )( #$$"( & ! * ) ) ,-( . ( /0"( 1$ ( ( 2+ 3" 4345514"$#14( . 6 & &* & * ( 2 2 & & * * & * & ( ! * & ( . 7 & * . * & & & & & * ( & & * & * 8 & ( ) & & * * ( . 4 4& * && * * && & * ( + * 9) ,9 ) - & & * : ( ) !"!#$% ,%- & 9) &!'$()* &! ,&- * & * ( && & * & * & ; ( ) % & * & && & * ( 2 8 & & * +, - & * 8 9) . / $ / * / $ * / ) %0/ / $ 12345 / , < ) 6 + #$$" 2++ 0540#1 2+ 3" 4345514"$#14 % %%% 4 / , %== (:(= > ? % %%% 4 /- looking carefully, a shepherd’s purse is blooming under the fence Bash List of papers This thesis is based on the following papers, which are referred to by their Roman numerals: I Slotte, T., Ceplitis, A., Neuffer, B., Hurka, H., and M. Lascoux. 2006. Intrageneric phylogeny of Capsella (Brassicaceae) and the -

Specialized and Rural Municipalities and Their Communities
Specialized and Rural Municipalities and Their Communities Updated December 18, 2020 Municipal Services Branch 17th Floor Commerce Place 10155 - 102 Street Edmonton, Alberta T5J 4L4 Phone: 780-427-2225 Fax: 780-420-1016 E-mail: [email protected] SPECIALIZED AND RURAL MUNICIPALITIES AND THEIR COMMUNITIES MUNICIPALITY COMMUNITIES COMMUNITY STATUS SPECIALIZED MUNICIPALITES Crowsnest Pass, Municipality of None Jasper, Municipality of None Lac La Biche County Beaver Lake Hamlet Hylo Hamlet Lac La Biche Hamlet Plamondon Hamlet Venice Hamlet Mackenzie County HIGH LEVEL Town RAINBOW LAKE Town Fort Vermilion Hamlet La Crete Hamlet Zama City Hamlet Strathcona County Antler Lake Hamlet Ardrossan Hamlet Collingwood Cove Hamlet Half Moon Lake Hamlet Hastings Lake Hamlet Josephburg Hamlet North Cooking Lake Hamlet Sherwood Park Hamlet South Cooking Lake Hamlet Wood Buffalo, Regional Municipality of Anzac Hamlet Conklin Hamlet Fort Chipewyan Hamlet Fort MacKay Hamlet Fort McMurray Hamlet December 18, 2020 Page 1 of 25 Gregoire Lake Estates Hamlet Janvier South Hamlet Saprae Creek Hamlet December 18, 2020 Page 2 of 25 MUNICIPALITY COMMUNITIES COMMUNITY STATUS MUNICIPAL DISTRICTS Acadia No. 34, M.D. of Acadia Valley Hamlet Athabasca County ATHABASCA Town BOYLE Village BONDISS Summer Village ISLAND LAKE SOUTH Summer Village ISLAND LAKE Summer Village MEWATHA BEACH Summer Village SOUTH BAPTISTE Summer Village SUNSET BEACH Summer Village WEST BAPTISTE Summer Village WHISPERING HILLS Summer Village Atmore Hamlet Breynat Hamlet Caslan Hamlet Colinton Hamlet -

Phylogenetic Position and Generic Limits of Arabidopsis (Brassicaceae)
PHYLOGENETIC POSITION Steve L. O'Kane, Jr.2 and Ihsan A. 3 AND GENERIC LIMITS OF Al-Shehbaz ARABIDOPSIS (BRASSICACEAE) BASED ON SEQUENCES OF NUCLEAR RIBOSOMAL DNA1 ABSTRACT The primary goals of this study were to assess the generic limits and monophyly of Arabidopsis and to investigate its relationships to related taxa in the family Brassicaceae. Sequences of the internal transcribed spacer region (ITS-1 and ITS-2) of nuclear ribosomal DNA, including 5.8S rDNA, were used in maximum parsimony analyses to construct phylogenetic trees. An attempt was made to include all species currently or recently included in Arabidopsis, as well as species suggested to be close relatives. Our ®ndings show that Arabidopsis, as traditionally recognized, is polyphyletic. The genus, as recircumscribed based on our results, (1) now includes species previously placed in Cardaminopsis and Hylandra as well as three species of Arabis and (2) excludes species now placed in Crucihimalaya, Beringia, Olimar- abidopsis, Pseudoarabidopsis, and Ianhedgea. Key words: Arabidopsis, Arabis, Beringia, Brassicaceae, Crucihimalaya, ITS phylogeny, Olimarabidopsis, Pseudoar- abidopsis. Arabidopsis thaliana (L.) Heynh. was ®rst rec- netic studies and has played a major role in un- ommended as a model plant for experimental ge- derstanding the various biological processes in netics over a half century ago (Laibach, 1943). In higher plants (see references in Somerville & Mey- recent years, many biologists worldwide have fo- erowitz, 2002). The intraspeci®c phylogeny of A. cused their research on this plant. As indicated by thaliana has been examined by Vander Zwan et al. Patrusky (1991), the widespread acceptance of A. (2000). Despite the acceptance of A. -

(Dr. Sc. Nat.) Vorgelegt Der Mathematisch-Naturwissenschaftl
Zurich Open Repository and Archive University of Zurich Main Library Strickhofstrasse 39 CH-8057 Zurich www.zora.uzh.ch Year: 2012 Flowers, sex, and diversity: Reproductive-ecological and macro-evolutionary aspects of floral variation in the Primrose family, Primulaceae de Vos, Jurriaan Michiel Posted at the Zurich Open Repository and Archive, University of Zurich ZORA URL: https://doi.org/10.5167/uzh-88785 Dissertation Originally published at: de Vos, Jurriaan Michiel. Flowers, sex, and diversity: Reproductive-ecological and macro-evolutionary aspects of floral variation in the Primrose family, Primulaceae. 2012, University of Zurich, Facultyof Science. FLOWERS, SEX, AND DIVERSITY. REPRODUCTIVE-ECOLOGICAL AND MACRO-EVOLUTIONARY ASPECTS OF FLORAL VARIATION IN THE PRIMROSE FAMILY, PRIMULACEAE Dissertation zur Erlangung der naturwissenschaftlichen Doktorwürde (Dr. sc. nat.) vorgelegt der Mathematisch-naturwissenschaftliche Fakultät der Universität Zürich von Jurriaan Michiel de Vos aus den Niederlanden Promotionskomitee Prof. Dr. Elena Conti (Vorsitz) Prof. Dr. Antony B. Wilson Dr. Colin E. Hughes Zürich, 2013 !!"#$"#%! "#$%&$%'! (! )*'+,,&$-+''*$.! /! '0$#1'2'! 3! "4+1%&5!26!!"#"$%&'(#)$*+,-)(*#! 77! "4+1%&5!226!-*#)$%.)(#!'&*#!/'%#+'.0*$)/)"$1'(12%-).'*3'0")"$*.)4&4'*#' "5*&,)(*#%$4'+(5"$.(3(-%)(*#'$%)".'(#'+%$6(#7.'2$(1$*.".! 89! "4+1%&5!2226!.1%&&'%#+',!&48'%'9,%#)()%)(5":'-*12%$%)(5"'"5%&,%)(*#'*3' )0"';."&3(#!'.4#+$*1"<'(#'0")"$*.)4&*,.'%#+'0*1*.)4&*,.'2$(1$*.".! 93! "4+1%&5!2:6!$"2$*+,-)(5"'(12&(-%)(*#.'*3'0"$=*!%14'(#'0*1*.)4&*,.' 2$(1$*.".>'5%$(%)(*#'+,$(#!'%#)0".(.'%#+'$"2$*+,-)(5"'%..,$%#-"'(#' %&2(#"'"#5($*#1"#).! 7;7! "4+1%&5!:6!204&*!"#")(-'%#%&4.(.'*3'!"#$%&''."-)(*#'!"#$%&''$"5"%&.' $%12%#)'#*#/1*#*204&4'%1*#!'1*2$0*&*!(-%&&4'+(.)(#-)'.2"-(".! 773! "4+1%&5!:26!-*#-&,+(#!'$"1%$=.! 7<(! +"=$#>?&@.&,&$%'! 7<9! "*552"*?*,!:2%+&! 7<3! !!"#$$%&'#""!&(! Es ist ein zentrales Ziel in der Evolutionsbiologie, die Muster der Vielfalt und die Prozesse, die sie erzeugen, zu verstehen. -

Taxonomic Key to the Saskatchewan Cryptantha Sarah Vinge-Mazer (Saskatchewan Conservation Data Centre) on Behalf of the Botanical Assessment Working Group
Taxonomic Key to the Saskatchewan Cryptantha Sarah Vinge-Mazer (Saskatchewan Conservation Data Centre) on behalf of the Botanical Assessment Working Group Introduction The Botanical Assessment Working Group has been working on placing Saskatchewan’s two perennial Cryptantha species since around 2011, following many questions and confusion from the local botanical community. The use of the name Cryptantha macounii was the source of much of the confusion, for which an explanation is now provided. The name Cryptantha macounii (=Oreocarya macounii) is a true synonym for Cryptantha celosioides (see Simpson). In Saskatchewan (and apparently Montana, see Lesica 2012) this name has instead been applied to Cryptantha spiculifera material. Furthering the confusion, the name Cryptantha nubigena has been applied to both C. spiculifera and C. celosioides in Saskatchewan and appears regularly in older taxonomic keys. The name C. macounii has to be maintained for C. spiculifera material in Sask. for now until the SKCDC can update its database with the name. C. spiculifera has been included as a synonym for C. macounii in the SKCDC vascular plant species list. This key was compiled from several sources and was written to clarify the placement of the Saskatchewan Cryptantha species and serve as interim information until the Flora of North America (FNA) on Boraginaceae is published. It is accurate to the best of our knowledge. Notes • Our two perennial species are retained in Cryptantha in this key but are expected to be moved under the genus Oreocarya in the FNA treatment (see Simpson) • The Manual of Montana Vascular Plants (Lesica 2012) provides species descriptions for all species that occur in Sask.