Movement, Habitat Use, and Food Habits Of
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Sauger Population Ecology in Three Missouri River Mainstem Reservoirs Brian D
South Dakota State University Open PRAIRIE: Open Public Research Access Institutional Repository and Information Exchange Electronic Theses and Dissertations 2006 Sauger Population Ecology in Three Missouri River Mainstem Reservoirs Brian D. S. Graeb South Dakota State University Follow this and additional works at: https://openprairie.sdstate.edu/etd Part of the Natural Resources and Conservation Commons Recommended Citation Graeb, Brian D. S., "Sauger Population Ecology in Three Missouri River Mainstem Reservoirs" (2006). Electronic Theses and Dissertations. 438. https://openprairie.sdstate.edu/etd/438 This Dissertation - Open Access is brought to you for free and open access by Open PRAIRIE: Open Public Research Access Institutional Repository and Information Exchange. It has been accepted for inclusion in Electronic Theses and Dissertations by an authorized administrator of Open PRAIRIE: Open Public Research Access Institutional Repository and Information Exchange. For more information, please contact [email protected]. Sauger Population Ecology in Three Missouri River Mainstem Reservoirs By Brian D. S. Graeb A dissertation submitted in partial fulfillment of the requirements for the degree Doctor of Philosophy Major in Biological Sciences South Dakota State University 2006 11 Sauger Population Ecology in Three Missouri River Mainstem Reservoirs This dissertation is approved as a credible and independent investigation by a candidate for the Doctor of Philosophy degree and is acceptable for meeting the dissertation requirements for this degree. Acceptance of this dissertation does not imply that the conclusions reached by the candidate are necessarily the conclusions of the major department. David W. Willis Date Dissertation Advisor Charles G. Scalet Date Head, Department of Wildlife and Fisheries Sciences iii Acknowledgements I would like to especially thank my wife, Stephanie for her support and guidance in all aspects of my life during my long tenure as a graduate student, and my advisor, Dr. -

Disconnect to Reconnect in Knox County Disconnect to Reconnect in Knox County
Disconnect to Reconnect in Knox County Disconnect to Reconnect in Knox County HistoryHistory of Knox Co. RecreationHistory WinnetoonKnox County was was founded organized in about by the 1892 Territorial and wasLegislature incorporated in 1857 KnoxCenter County was provides platted many in 1901. beautiful It was parks named where for its you can play asand a villagenamed inL’Eau 1900. Qui It Court, was named meaning after in English, the Wisconsin “Running farm Water.” of volleyball,location have near a picnic, the geographical or walk through center an arboretum.of Knox Most area anThe acquaintance name was changedof a railroad to Knox official. by Aa poststatute office passed has Februarybeen in townsCounty. have community Center was swimming incorporated pools asas well,a village so stop and relax operation21, 1873, in which Winnetoon took effect since April 1891. 1, 1873. In 1996 Chief Standing poolside.in 1904 and is the county seat of Knox County, Bear Bridge was completed, bridging the shores of Knox County, Nebraska. near Niobrara, with South Dakota across the Missouri River. Knox Water Sports County containsParks about 1,050& Recreation miles, or 672,000 acres. If you enjoy boating, fishing, or swimming, you’ve found the ultimate WINNETOON MALL & POST OFFICE - Antiques, woodworking, play area.Parks Whether &you’re Recreation fishing in the Lewis and Clark Lake, furniture refinishing & repair. Natural Food Coop also. Visit Privy Missouri River, or the Niobrara River, you will find a wide variety of Path and the garden areas. Go back in time! fish includingCENTER walleye, PARK smallmouth- A quaint, bass, little largemouth park with bass,a northern Attractions playground, picnic shelter and restroom in the Explore an area filled with culture, history, recreation and adventure. -

Fisheries Management Plan
What Makes Lewis and Clark Lake and the Missouri River Fisheries Special Fisheries Management Plan This section of the Missouri River and Lake Lewis and for Lewis and Clark Lake and the Missouri River Upstream Clark has a rich diversity of habitats, sport fish and native riverine species. This diversity makes the area one of the and Downstream of the Reservoir 2017 - 2021 last places on the Missouri River where vestiges of the This is a summary of the Fisheries Management Plan for Lewis and Clark Lake and the Missouri River Upstream historical habitat remain. This area also is the hot bed of and Downstream of the Reservoir (Lewis and Clark Study Area). It is part of the overall Missouri River Fisheries AIS infestations in the state, with Asian carp thriving below Strategic Management Plan. Gavins Point Dam and zebra mussels infesting Lewis and Clark Lake. We are challenged to prevent the spread of Major Issues Identified for Lewis and Clark Lake and the Missouri River Upstream and Downstream of the Reservoir: these species and to find ways to mitigate their impacts to • Shoreline and boat access on can be limited due to a variety of factors. native fauna. Lewis and Clark Lake also is home to South • Habitat quantity and quality negatively impact fish populations and fishing opportunity. Dakota’s busiest state park and is a hub for water recreation • Many knowledge gaps exist for fish population dynamics. activities. South Dakota shares management responsibilities • New and established aquatic invasive species could potentially impact the fishery and recreation. of the area downstream of the Nebraska state-line to the • Government and public interactions related to management can be challenging. -

Foundation Document Missouri National Recreational River Nebraska and South Dakota August 2017
NATIONAL PARK SERVICE • U.S. DEPARTMENT OF THE INTERIOR Foundation Document Missouri National Recreational River Nebraska and South Dakota August 2017 MNRR_FD_PRINT.indd 1 9/22/17 11:20 AM To 90 and Chamberlain To 90 To 90 and Mitchell To 90 To 90 and Sioux Falls Foundation Document 18 18 North 50 0 5 10 Kilometers 281 J a 0 5 10 Miles s m e e d s n A R Lake iv Missouri National Campground Canoe access e r Andes e Recreational River Lake Franc Lak is 37 C a 281 18 50 Unpaved road Primitive campsite Boat launch s e 81 NORTH POINT STATE YANKTON SIOUX RECREATION AREA Lewis and Clark Hiking trail Marina INDIAN RESERVATION National Historic Trail driving route Fort Randall Dam 46 46 50 Visitor Center Fort Pickstown 39-MILE DISTRICTWagner Randall Missouri National Recreational RiverC (Information summer only) Dam h Spillway o te a SOUTH DAKOTA ek Overlook u 59-MILE DISTRICT re Missouri National Recreational River To Winner C C l Fort Randall r 18 l e 29 SOUTH DAKOTA a e Historic Site r d k e NEBRASKA n v a 50 i KARL Avon R R Tyndall k Marty e MUNDT O re 12 ya C NATIONAL te Tr 19 11 ail le WILDLIFE u r x Po REFUGE 37 B u nc Tabor io a Yankton Sioux S C re ek Old Baldy Treaty Monument 281 Gavins Point National Fish 12 Overlook N Hatchery and Aquarium Yankton 11 Sunshine a g Butte tive Missouri National Recreational River Headquarters i Bottom A B me SPRINGFIELD STATE 52 50 Historic Meridian Bridge rica n S RECREATION AREA LEWIS AND CLARK STATE Riverside Park cen 0 ic RECREATION AREA M By Springfield 52 SPIRIT MOUND Spencer ISS wa James River IOWA OU y HISTORIC PRAIRIE Ni R Gavins Island ob I Le Lake ra R w Point 48 ra IV Santee is a ake n R Lynch E nd Clark L Dam 121 St. -

Food Habits of Bigmouth and Smallmouth Buffalo in Lewis and Clark Lake and the Missouri River Thomas S
South Dakota State University Open PRAIRIE: Open Public Research Access Institutional Repository and Information Exchange Theses and Dissertations 1964 Food Habits of Bigmouth and Smallmouth Buffalo in Lewis and Clark Lake and the Missouri River Thomas S. McComish Follow this and additional works at: http://openprairie.sdstate.edu/etd Part of the Natural Resources and Conservation Commons Recommended Citation McComish, Thomas S., "Food Habits of Bigmouth and Smallmouth Buffalo in Lewis and Clark Lake and the Missouri River" (1964). Theses and Dissertations. 175. http://openprairie.sdstate.edu/etd/175 This Thesis - Open Access is brought to you for free and open access by Open PRAIRIE: Open Public Research Access Institutional Repository and Information Exchange. It has been accepted for inclusion in Theses and Dissertations by an authorized administrator of Open PRAIRIE: Open Public Research Access Institutional Repository and Information Exchange. For more information, please contact [email protected]. FOCD I!ABITS CF BIG!11>UTD AND SHt..LU:OUTH BUFFALO MISSOUi?.I RIVER BY TllOHAS S. McCOMISll A th�ois suboittcd i::.i. ?��tic.1 folf illr:-.cnt of the rcqui'l:ctr.cnts for the ec::;rcc �-::Jotc:: oi Science, Dcl)n.:tr.:ent of tliltlli:fo E�i.�scr..cnt, South D�kota State Collc�c of Agriculture �nd }:iechanic Arts 1964 e- ""' :TW T'\A lttiT.t. <:;Tti. TF UNIVERSITY LIBRARY · Z·:::::. t!-..e:.i::: is �?;?i:ovcci as a. creditable and independent i:-ave:stication by :i ca:-..di�c.::c ·ror the degi·ec, 1'1aoter of Science, and i::; .:iccc?'..:��:c �::; ::1ec::i:.1;3 t:!c t!1csis requirements for this degree, a'!:� .1ccess�n:�ly the conclu:::ious of the r.�jor depart� iii :;: wish. -

Bureau of Sport Fisheries and Wildlife and Possibly Flathead Catfish Have
24 Technical Paper 56: Bureau of Sport Fisheries and Wildlife and possibly flathead catfish have increased; walleye, and sauger was faster than in Oahe the abundance of most other species for which Reservoir. Growth estimates in Oahe Reser we have reliable data has declined. A similar voir were based only on the early years of change has occurred in Oahe Reservoir, where impoundment, when growth rates are generally catfish, white bass, and walleye populations the highest. have increased (Higham, 1968). The following species, though most were never present in Growth of northern pike, river carpsucker, large numbers, have become rare in Lake largemouth bass, walleye, and sauger was Francis Case since the first years of impound better than in most other waters in this latitude ment: flathead chub, stonecat, black bullhead, in the United States, but carp and drum growth largemouth bass, green sunfish, orangespotted was poorer. sunfish, and bluegill. The sport-fishing catch rate in Lake Francis The main forage species in the reservoir Case declined from 1954 to 1960, although the were gizzard shad, emerald shiner, and yellow catch rates for sauger and crappie increased perch. These species did not appear to be during that period. present in large enough numbers to provide an abundant forage fish population as reported The sauger population declined between in many lakes or reservoirs with shad and 1956 and 1962, while the walleye population yellow perch. As Selgeby (1968) suggests, the increased from 1959 to 1962 (fig. 8). By 1967 high zooplankton standing crop and the com the sauger population, which was previously paratively large individual size of the larger than that of walleye, had become the cladocerans may indicate an open niche in the smaller of the two. -

WALLEYE Stizostedion V
FIR/S119 FAO Fisheries Synopsis No. 119 Stizostedion v. vitreum 1,70(14)015,01 SYNOPSIS OF BIOLOGICAL DATA ON THE WALLEYE Stizostedion v. vitreum (Mitchill 1818) A, F - O FOOD AND AGRICULTURE ORGANIZATION OFTHE UNID NP.TION3 FISHERIES SYNOR.9ES This series of documents, issued by FAO, CSI RO, I NP and NMFS, contains comprehensive reviews of present knowledge on species and stocks of aquatic organisms of present or potential economic interest. The Fishery Resources and Environment Division of FAO is responsible for the overall coordination of the series. The primary purpose of this series is to make existing information readily available to fishery scientists according to a standard pattern, and by so doing also to draw attention to gaps in knowledge. It is hoped 41E11 synopses in this series will be useful to other scientists initiating investigations of the species concerned or or rMaIeci onPs, as a means of exchange of knowledge among those already working on the species, and as the basis íoi study of fisheries resources. They will be brought up to date from time to time as further inform.'t:i available. The documents of this Series are issued under the following titles: Symbol FAO Fisheries Synopsis No. 9R/S CS1RO Fisheries Synopsis No. INP Sinopsis sobre la Pesca No. NMFS Fisheries Synopsis No. filMFR/S Synopses in these series are compiled according to a standard outline described in Fib/S1 Rev. 1 (1965). FAO, CSI RO, INP and NMFS are working to secure the cooperation of other organizations and of individual scientists in drafting synopses on species about which they have knowledge, and welcome offers of help in this task. -

Large-Scale Impacts of Hydroelectric Development
27 Large-scale impacts of hydroelectric development D.M. Rosenberg,1 F. Berkes, R.A. Bodaly, R.E. Hecky, C.A. Kelly, and J.W.M. Rudd Abstract: The substantial size of some hydroelectric projects and the extensive total surface area covered by reservoirs globally require that research determining the impacts of these developments be done at ever-increasing spatial and temporal scales. As a consequence of this research, new views are emerging about the spatial extent and longevity of the environmental and social impacts of such developments. New findings challenge the notion of hydroelectric development as a benign alternative to other forms of power generation. This review examines the intertwined environmental and social effects of methylmercury bioaccumulation in the food web, emission of greenhouse gases from reservoirs, downstream effects of altered flows, and impacts on biodiversity, each of which operates at its own unique spatial and temporal scales. Methylmercury bioaccumulation occurs at the smallest spatial and temporal scales of the four impacts reviewed, whereas downstream effects usually occur at the largest scales. Greenhouse gas emissions, the newest surprise connected with large-scale hydroelectric development, are relatively short term but eventually may have important global-scale consequences. Limitation of biodiversity by hydroelectric development usually occurs at intermediate spatial and temporal scales. Knowledge developed from working at expanded spatial and temporal scales should be an important part of future decision making for large-scale hydroelectric development. Key words: hydroelectric development, large-scale, environmental impacts, social impacts. Résumé : La dimension considérable de certains projets hydroélectriques et les vastes surfaces totales globalement couvertes par les réservoirs nécessitent que la recherche menée pour déterminer les impacts de ces développements soit conduite à des échelles d’espace et de temps de plus en plus grandes. -

Missouri River Gavins Point to Ponca State Park
MISSOURI RIVER GAVINS POINT TO PONCA STATE PARK Recommendation For Inclusion Under Section S(d) of the Wild and Scenic Rivers Act Department of the Interior Bureau of Outdoor Recreation AUGUST 1971 MISSOURI RIVER -- GAVINS POINT DAM TO PONCA STATE PARK I. INTRODUCTION Beginning at Three Forks, Montana, the Missouri River flows south easterly over 2,300 miles to join the Mississippi a few miles north of St. Louis, Missouri. It ranks as the second longest and, his torically, one of the most famous rivers in the United States. It was the primary route of Lewis and Clark on their epic journey of exploration. Long before the advent of transcontinental railways, it shared with the Oregon Trail and the Santa Fe Trail the distinc tion of being one of the three main thoroughfares to the Far West, witnessing a cavalcade of fur trappers, missionaries, gold miners, and pioneer settlers. But here, instead of covered wagons, convey ance was by pirogue, keelboat and steamboat. It was a wild and un predictable river that carried millions of pounds of freight. Waters of this mighty river that for centuries ran uncontrolled are now harnessed in the upper and middle reaches by a series of multi purpose dams and reservoirs. In its lower reaches, the Missouri has been further tamed by channelization. The net effect of man's endeavors has been improved flood control and navigation, increased hydroelectric power generation, irrigation water and creation of new opportunities for outdoor recreation. However~ these worthy projects have also eliminated most of the extraordinary aesthetic, historic and wildlife values associated with the river in its natural condition. -
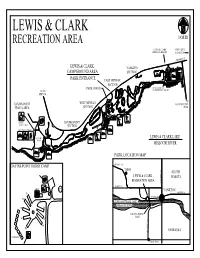
W:\BASEMAPS\Dist190\Campground Maps\Campground Maps 2007
LEWIS & CLARK RECREATION AREA NORTH LEWIS & CLARK 4 MILES TO MARINA & RESORT YANKTON SD HWY 52 LEWIS & CLARK YANKTON CAMPGROUND AREA SECTION PARK ENTRANCE EAST MIDWAY SECTION PARK OFFICE RESTAURANT & TO SD CONVENIENCE STORE HWY 50 TO GAVINS POINT WEST MIDWAY GAVINS POINT TRAILS AREA SECTION DAM HORSE GAVINS POINT TRAIL CAMP SECTION MULTI-USE TRAIL NATURE LEWIS & CLARK LAKE TRAIL MISSOURI RIVER PARK LOCATION MAP GAVINS POINT HORSE CAMP SD HWY 50 TABOR SOUTH LEWIS & CLARK DAKOTA RECREATION AREA US HWY 81 SD HWY 52 HC 1 VT YANKTON SD HWY 50 H1 #1 W SD HWY 52 H2 LEWIS AND CLARK LAKE MISSOURI RIVER H3 H8 W GAVINS POINT DAM H4 H7 H6 H5 NEBRASKA NB HWY 121 CORRALS CROFTON Map Legend Below are the symbols found on the campground maps for the Accessible South Dakota state parks. Use this guide to identify the features Facilities found in each campground. Campground Amphitheater Swim Beach Group Archery Ski Camping Area Trail Beach Entrance Bicycle Boat Station Trail Ramp Comfort Cross-Country Canoeing Station Ski Trail Dump Sledding Area Accessible Station Fishing Dock Fish Cleaning Disc Playground Station Golf Fish Cleaning Hiking Picnic Table Trail Area Telephone Horse Trail/ Picnic Horse Camp Shelter Resort/ Concessionaire Double-up Sites Electrical If two camping units share a site, each unit must pay the daily E Campsite camping fee. A camping unit is a vehicle, motor home, camping bus, pull-type camper, tent or any other device designed for * 50 amp electrical sleeping. Additional tents for a family (parent or grandparents campsite all others 30 amp and their unmarried minor children) are considered a single Vault camping unit. -

Waterfowl Guide
2018 WATERFOWL GUIDE • Early Teal • Duck and Goose • Youth • Light Goose Conservation Order INFORMATION FOR: INFORMATION OutdoorNebraska.org Paid Advertisement TABLE OF CONTENTS ABOUT THIS GUIDE Early Teal ���������������������������������������������������� 8 The Waterfowl Guide is a publication of the Nebraska Game and Parks Commission. It is a summary of Duck and Coot . .11 Nebraska’s regulations, orders and statutes pertaining Youth Waterfowl �����������������������������������������11 to waterfowl hunting and is not intended to be a Dark Goose ������������������������������������������������� 15 complete digest of regulations, orders and statutes. A complete listing may be found at the following links: White-fronted Goose ���������������������������������� 16 Regulations and Orders: Light Goose Regular Season . 16 http://outdoornebraska.org/regulations/ Light Goose Conservation Order ���������������� 16 Statutes: Conservation Officers. 23 http://law.justia.com/codes/nebraska/2016/ Sunrise/Sunset Schedule . 24 chapter-37/ Duck Identification ��������������������������������26-27 U.S. Fish and Wildlife Service Regulations: fws.gov/hunting Zone Maps: ����������������������������������������������������� Paid advertisements included in this publication do not Teal ��������������������������������������������������������� 8 represent an endorsement by the Commission for the Duck and Coot. 12-14 products, services or claims made by the advertisers. Dark Goose . 15 The Commission reserves the right to reject any advertising for any reason. Light Goose ������������������������������������������� 17 NEBRASKA GAME AND PARKS COMMISSION OFFICES HEADQUARTERS/SOUTHEAST District 2200 N. 33rd St. Lincoln, NE 68503-0370 402-471-0641 SOUTHWEST District 301 E. State Farm Road North Platte, NE 69101-0430 308-535-8025 NORTHEAST District 2201 N. 13th St. Norfolk, NE 68701-2267 402-370-3374 NORTHWEST District Kearney Service Center Bassett Service Center Omaha Service Center 299 Husker Road 1617 First Ave. 524 Panzer St. -

Reconnaissance Survey of Lewis and Clark on the Missouri National Recreational River, Nebraska and South Dakota
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Papers in Natural Resources Natural Resources, School of 12-1999 Reconnaissance Survey of Lewis and Clark on the Missouri National Recreational River, Nebraska and South Dakota Anne P. Diffendal University of Nebraska-Lincoln, [email protected] Gary E. Moulton University of Nebraska-Lincoln, [email protected] Michael Shambaugh-Miller University of Nebraska - Lincoln Robert F. Diffendal University of Nebraska - Lincoln, [email protected] Follow this and additional works at: https://digitalcommons.unl.edu/natrespapers Part of the Natural Resources and Conservation Commons Diffendal, Anne P.; Moulton, Gary E.; Shambaugh-Miller, Michael; and Diffendal, Robert F., "Reconnaissance Survey of Lewis and Clark on the Missouri National Recreational River, Nebraska and South Dakota" (1999). Papers in Natural Resources. 294. https://digitalcommons.unl.edu/natrespapers/294 This Article is brought to you for free and open access by the Natural Resources, School of at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Papers in Natural Resources by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. - - Reconnaissance Survey of Lewis and Clark -,- on the Missouri National Recreational River, Nebraska and South Dakota Decem her 1999 n - .- Reconnaissance Survey of Lewis and Clark - on the - Missouri National Recreational River, Nebraska and South Dakota by Anne P. Diffendal and Gary E. Moulton - - - - Cartography by Michael Shambaugh-Miller - Geology by Robert F. Diffendal - - - - - December 1999 - - Contract No. 1443-CX-1505-9-0011 Research Passage of Lewis and Clark Acknowledgments - The authors are grateful for the assistance of Duane R. Molman, Conservation and Survey Division, University of Nebraska-Lincoln; Laura Banker, Omaha District Office, U.S.