(Anthonomus Signatus) on Raspberry (Rubus Idaeus) in Maine Christina S
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Integration of Entomopathogenic Fungi Into IPM Programs: Studies Involving Weevils (Coleoptera: Curculionoidea) Affecting Horticultural Crops
insects Review Integration of Entomopathogenic Fungi into IPM Programs: Studies Involving Weevils (Coleoptera: Curculionoidea) Affecting Horticultural Crops Kim Khuy Khun 1,2,* , Bree A. L. Wilson 2, Mark M. Stevens 3,4, Ruth K. Huwer 5 and Gavin J. Ash 2 1 Faculty of Agronomy, Royal University of Agriculture, P.O. Box 2696, Dangkor District, Phnom Penh, Cambodia 2 Centre for Crop Health, Institute for Life Sciences and the Environment, University of Southern Queensland, Toowoomba, Queensland 4350, Australia; [email protected] (B.A.L.W.); [email protected] (G.J.A.) 3 NSW Department of Primary Industries, Yanco Agricultural Institute, Yanco, New South Wales 2703, Australia; [email protected] 4 Graham Centre for Agricultural Innovation (NSW Department of Primary Industries and Charles Sturt University), Wagga Wagga, New South Wales 2650, Australia 5 NSW Department of Primary Industries, Wollongbar Primary Industries Institute, Wollongbar, New South Wales 2477, Australia; [email protected] * Correspondence: [email protected] or [email protected]; Tel.: +61-46-9731208 Received: 7 September 2020; Accepted: 21 September 2020; Published: 25 September 2020 Simple Summary: Horticultural crops are vulnerable to attack by many different weevil species. Fungal entomopathogens provide an attractive alternative to synthetic insecticides for weevil control because they pose a lesser risk to human health and the environment. This review summarises the available data on the performance of these entomopathogens when used against weevils in horticultural crops. We integrate these data with information on weevil biology, grouping species based on how their developmental stages utilise habitats in or on their hostplants, or in the soil. -
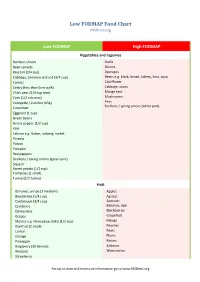
Low FODMAP Food Chart Ibsdiets.Org
Low FODMAP Food Chart IBSDiets.org Low FODMAP High FODMAP Vegetables and Legumes Bamboo shoots Garlic Bean sprouts Onions Broccoli (3/4 cup) Aparagus Cabbage, common and red (3/4 cup) Beans e.g. black, broad, kidney, lima, soya Carrots Cauliflower Celery (less than 5cm stalk) Cabbage, savoy Chick peas (1/4 cup max) Mange tout Corn (1/2 cob max) Mushrooms Courgette / Zucchini (65g) Peas Scallions / spring onions (white part) Cucumber Eggplant (1 cup) Green beans Green pepper (1/2 cup) Kale Lettuce e.g. Butter, iceberg, rocket Parsnip Potato Pumpkin Red peppers Scallions / spring onions (green part) Squash Sweet potato (1/2 cup) Tomatoes (1 small) Turnip (1/2 turnip) Fruit Bananas, unripe (1 medium) Apples Blueberries (1/4 cup) Apricot Cantaloupe (3/4 cup) Avocado Cranberry Bananas, ripe Clementine Blackberries Grapes Grapefruit Melons e.g. Honeydew, Galia (1/2 cup) Mango Kiwifruit (2 small) Peaches Lemon Pears Orange Plums Pineapple Raisins Raspberry (30 berries) Sultanas Rhubarb Watermelon Strawberry For up to date and extensive information go to www.IBSDiets.org Meat and Substitutes Beef Sausages (check ingredients) Chicken Processed meat (check ingredients) Lamb Pork Quorn mince Cold cuts e.g. Ham and turkey breast Breads, Cereals, Grains and Pasta Oats Barley Quinoa Bran Gluten free foods e.g. breads, pasta Cous cous Savory biscuits Gnocchi Buckwheat Granola Chips / crisps (plain) Muesli Cornflour Muffins Oatmeal (1/2 cup max) Rye Popcorn Semolina Pretzels Spelt Rice e.g. Basmati, brown, white Wheat foods e.g. Bread, cereal, pasta -

Anthonomus Eugenii Pepper Weevil
Pest specific plant health response plan: Outbreaks of Anthonomus eugenii Figure 1. Adult Anthonomus eugenii. © Fera Science Ltd 1 © Crown copyright 2020 You may re-use this information (not including logos) free of charge in any format or medium, under the terms of the Open Government Licence. To view this licence, visit www.nationalarchives.gov.uk/doc/open-government-licence/ or write to the Information Policy Team, The National Archives, Kew, London TW9 4DU, or e-mail: [email protected] This document is also available on our website at: https://planthealthportal.defra.gov.uk/pests-and-diseases/contingency-planning/ Any enquiries regarding this document should be sent to us at: The UK Chief Plant Health Officer Department for Environment, Food and Rural Affairs Room 11G32 National Agri-Food Innovation Campus Sand Hutton York YO41 1LZ Email: [email protected] 2 Contents 1. Introduction and scope ......................................................................................................... 4 2. Summary of threat................................................................................................................. 4 3. Risk assessments ................................................................................................................. 5 4. Actions to prevent outbreaks ............................................................................................... 5 5. Response .............................................................................................................................. -

IOBC/WPRS Working Group “Integrated Plant Protection in Fruit
IOBC/WPRS Working Group “Integrated Plant Protection in Fruit Crops” Subgroup “Soft Fruits” Proceedings of Workshop on Integrated Soft Fruit Production East Malling (United Kingdom) 24-27 September 2007 Editors Ch. Linder & J.V. Cross IOBC/WPRS Bulletin Bulletin OILB/SROP Vol. 39, 2008 The content of the contributions is in the responsibility of the authors The IOBC/WPRS Bulletin is published by the International Organization for Biological and Integrated Control of Noxious Animals and Plants, West Palearctic Regional Section (IOBC/WPRS) Le Bulletin OILB/SROP est publié par l‘Organisation Internationale de Lutte Biologique et Intégrée contre les Animaux et les Plantes Nuisibles, section Regionale Ouest Paléarctique (OILB/SROP) Copyright: IOBC/WPRS 2008 The Publication Commission of the IOBC/WPRS: Horst Bathon Luc Tirry Julius Kuehn Institute (JKI), Federal University of Gent Research Centre for Cultivated Plants Laboratory of Agrozoology Institute for Biological Control Department of Crop Protection Heinrichstr. 243 Coupure Links 653 D-64287 Darmstadt (Germany) B-9000 Gent (Belgium) Tel +49 6151 407-225, Fax +49 6151 407-290 Tel +32-9-2646152, Fax +32-9-2646239 e-mail: [email protected] e-mail: [email protected] Address General Secretariat: Dr. Philippe C. Nicot INRA – Unité de Pathologie Végétale Domaine St Maurice - B.P. 94 F-84143 Montfavet Cedex (France) ISBN 978-92-9067-213-5 http://www.iobc-wprs.org Integrated Plant Protection in Soft Fruits IOBC/wprs Bulletin 39, 2008 Contents Development of semiochemical attractants, lures and traps for raspberry beetle, Byturus tomentosus at SCRI; from fundamental chemical ecology to testing IPM tools with growers. -

Artículos Originales
BOLETIN DEL MUSEO ENTOMOLÓGICO FRANCISCO LUÍS GALLEGO ARTÍCULOS ORIGINALES SPECIES OF THE BEETLE GENUS ANTHONOMUS GERMAR, 1817 (CURCULIONIDAE: CURCULIONINAE: ANTHONOMINI) OF QUARANTINE IMPORTANCE INTERCEPTED AT U.S. PORTS OF ENTRY Allan H. Smith-Pardo. Entomologist. USDA-APHIS-PPQ, 389 Oyster Point Blvd., Suite 2, South San Francisco, CA. 94080. United States of America. [email protected] Abstract This paper presents a discussion on the taxonomy of weevils of the genus Anthonomus and presents diagnostic characters useful for the identification of adult and immature stages of the genus. In addition, I ran a search in the USDA’s AQAS database for interceptions of species of Anthonomus of quarantine importance that have been intercepted at United States ports of entry. In total, six species of quarantine importance have been intercepted: Anthonomus flavus, Anthonomus grandis, Anthonomus melanosticus, Anthonomus pomorum, Anthonomus rubi, and Anthonomus sisyphus. Photographs and information on the origin and hosts of these interceptions are included. Key words Weevil, pest, crop, imported commodities, diagnostic character Resumen En este artículo se presenta una discusión acerca de la taxonomía de escarabajos- picudos del genero Anthonomus y se presentan los caracteres diagnósticos para la identificación de estadios inmaduros y de adultos. Adicionalmente, se hizo una búsqueda de las intercepciones de especies del genero Anthonomus de 7 Volumen 7 • Número 1 Marzo • 2015 importancia cuarentenaria en los EEUU plants of the family Rosaceae and is an que han sido interceptados en puertos de important pest of strawberry (Fragaria x entrada. En total se han interceptado seis ananassa) and raspberry (Rubus idaeus); especies de importancia cuarentenaria and the pepper weevil, Anthonomus de acuerdo a la base de datos AQAS del eugenii, which feeds on plants of the USDA: Anthonomus flavus, Anthonomus genus Capsicum and Solanum] or as grandis, Anthonomus melanosticus, biological control agents of invasive Anthonomus pomorum, Anthonomus rubi plants [e.g. -

Growing Raspberries & Blackberries
Growing Raspberries Blackberries& in the Inland Northwest & Intermountain West by Danny L. Barney, Michael Colt, Jo Ann Robbins, and Maurice Wiese The authors Danny Barney, Extension Horticulturist and Superinten- dent of the University of Idaho Sandpoint Research & Extension Center Michael Colt, Extension Horticulturist at the UI Parma Research and Extension Center Jo Ann Robbins, Extension Educator in Blaine County Maurice Wiese, Plant Pathologist, UI Department of Plant, Soil, and Entomological Sciences © 1999 University of Idaho Contents How raspberries and blackberries grow ......... 3 Selecting and preparing your site................... 5 Designing your plots ..................................... 6 Plant and row spacing ............................. 6 Support................................................... 8 Trellis posts ........................................... 12 Selecting your plants .................................. 14 Planting ...................................................... 14 Caring for your berries ................................ 15 Irrigation............................................... 15 Fertilization........................................... 19 Weed control ........................................ 22 Winter protection ................................. 23 Pruning................................................. 24 Harvesting and storing your berries ........................................... 27 Controlling insect pests and diseases ........... 28 Insects and mites ................................... 28 -

Phenolics in Human Health
International Journal of Chemical Engineering and Applications, Vol. 5, No. 5, October 2014 Phenolics in Human Health T. Ozcan, A. Akpinar-Bayizit, L. Yilmaz-Ersan, and B. Delikanli with proteins. The high antioxidant capacity makes Abstract—Recent research focuses on health benefits of polyphenols as an important key factor which is involved in phytochemicals, especially antioxidant and antimicrobial the chemical defense of plants against pathogens and properties of phenolic compounds, which is known to exert predators and in plant-plant interferences [9]. preventive activity against infectious and degenerative diseases, inflammation and allergies via antioxidant, antimicrobial and proteins/enzymes neutralization/modulation mechanisms. Phenolic compounds are reactive metabolites in a wide range of plant-derived foods and mainly divided in four groups: phenolic acids, flavonoids, stilbenes and tannins. They work as terminators of free radicals and chelators of metal ions that are capable of catalyzing lipid oxidation. Therefore, this review examines the functional properties of phenolics. Index Terms—Health, functional, phenolic compounds. I. INTRODUCTION In recent years, fruits and vegetables receive considerable interest depending on type, number, and mode of action of the different components, so called as “phytochemicals”, for their presumed role in the prevention of various chronic diseases including cancers and cardiovascular diseases. Plants are rich sources of functional dietary micronutrients, fibers and phytochemicals, such -

(Coleoptera) from European Eocene Ambers
geosciences Review A Review of the Curculionoidea (Coleoptera) from European Eocene Ambers Andrei A. Legalov 1,2 1 Institute of Systematics and Ecology of Animals, Siberian Branch, Russian Academy of Sciences, Frunze Street 11, 630091 Novosibirsk, Russia; [email protected]; Tel.: +7-9139471413 2 Biological Institute, Tomsk State University, Lenina Prospekt 36, 634050 Tomsk, Russia Received: 16 October 2019; Accepted: 23 December 2019; Published: 30 December 2019 Abstract: All 142 known species of Curculionoidea in Eocene amber are documented, including one species of Nemonychidae, 16 species of Anthribidae, six species of Belidae, 10 species of Rhynchitidae, 13 species of Brentidae, 70 species of Curcuionidae, two species of Platypodidae, and 24 species of Scolytidae. Oise amber has eight species, Baltic amber has 118 species, and Rovno amber has 16 species. Nine new genera and 18 new species are described from Baltic amber. Four new synonyms are noted: Palaeometrioxena Legalov, 2012, syn. nov. is synonymous with Archimetrioxena Voss, 1953; Paleopissodes weigangae Ulke, 1947, syn. nov. is synonymous with Electrotribus theryi Hustache, 1942; Electrotribus erectosquamata Rheinheimer, 2007, syn. nov. is synonymous with Succinostyphlus mroczkowskii Kuska, 1996; Protonaupactus Zherikhin, 1971, syn. nov. is synonymous with Paonaupactus Voss, 1953. Keys for Eocene amber Curculionoidea are given. There are the first records of Aedemonini and Camarotini, and genera Limalophus and Cenocephalus in Baltic amber. Keywords: Coleoptera; Curculionoidea; fossil weevil; new taxa; keys; Palaeogene 1. Introduction The Curculionoidea are one of the largest and most diverse groups of beetles, including more than 62,000 species [1] comprising 11 families [2,3]. They have a complex morphological structure [2–7], ecological confinement, and diverse trophic links [1], which makes them a convenient group for characterizing modern and fossil biocenoses. -

Report on Beetles (Coleoptera) Collected from the Dartington Hall Estate, 2013 by Dr Martin Luff
Report on beetles (Coleoptera) collected from the Dartington Hall Estate, 2013 by Dr Martin Luff. 1. Introduction This year has been a particularly busy one for my work on the beetles of the Estate. I recorded numbers of species on 11 separate dates from April (at the end of the cold spring) through to mid- November. The generally warm and dry summer enabled me to record much more by sweeping herbaceous vegetation around field margins, especially in Hill Park. At the end of May I was assisted by an old friend of mine, and former fellow student, Dr Colin Welch (RCW), who is an authority on the Staphylinidae (rove beetles). I was also provided with the contents of the nest boxes from Dartington Hills in February and September, thanks to Will Wallis and Mike Newby. Finally Mary Bartlett again encouraged me to examine the fauna of her compost heap in November. 2. Results A total of 201 beetle species from 35 families were recorded. This is considerably more than in any previous year that I have collected at Dartington. Of these, 85 species were not recorded in my earlier lists (Luff, 2010-12). The overall number of species that I have recorded from the Estate is now 369, which is almost 10% of the entire British beetle fauna. The bird nest boxes yielded 13 species, with over half being new to the Estate, despite having examined the boxes in previous years; the contents of the boxes were also rather different between spring and autumn, with only four species common to both. -

Raspberry Pomace - Composition, Properties and Application
View metadata, citation and similar papers at core.ac.uk brought to you by CORE European Journal ISSN 2449-8955 Review Article of Biological Research Raspberry pomace - composition, properties and application Agnieszka Joanna Brodowska Institute of General Food Chemistry, Faculty of Biotechnology and Food Sciences, Łódź University of Technology, Stefanowskiego 4/10, 90-924 Łódź, Poland * Corresponding author: Agnieszka Brodowska; E-mail: [email protected] Received: 19 February 2017; Revised submission: 24 March 2017; Accepted: 03 April 2017 Copyright: © The Author(s) 2017. European Journal of Biological Research © T.M.Karpi ński 2017. This is an open access article licensed under the terms of the Creative Commons Attribution Non-Commercial 4.0 International License, which permits unrestricted, non-commercial use, distribution and reproduction in any medium, provided the work is properly cited. DOI : http://dx.doi.org/10.5281/zenodo.495190 ABSTRACT industry by-product, is considered as a potential food ingredient. Its pomace contains plenty of Raspberry pomace can be valorised due to its valuable components such as carbohydrates, nutritionally favourable effect on human health. proteins, fats, fibre, flavours, pectins, vitamins, It is an important source of polyphenols, ellagic similar to the composition of whole raspberries [2]. acid, ellagitannins, tocopherols, unsaturated fatty Moreover, raspberry pomace is rich in a large group acids, and dietary fibre. Thus, raspberry pomace of various phenolics especially ellagitannins, can be considered as a potential raw material to proanthocyanidins, anthocyanins, flavonols, and receive products rich in polyphenols or dietary phenolic acids (especially, ellagic acid) which are fibre, which can provide healthy properties to food also predominant in berries [3]. -

Chapter 1 Definitions and Classifications for Fruit and Vegetables
Chapter 1 Definitions and classifications for fruit and vegetables In the broadest sense, the botani- Botanical and culinary cal term vegetable refers to any plant, definitions edible or not, including trees, bushes, vines and vascular plants, and Botanical definitions distinguishes plant material from ani- Broadly, the botanical term fruit refers mal material and from inorganic to the mature ovary of a plant, matter. There are two slightly different including its seeds, covering and botanical definitions for the term any closely connected tissue, without vegetable as it relates to food. any consideration of whether these According to one, a vegetable is a are edible. As related to food, the plant cultivated for its edible part(s); IT botanical term fruit refers to the edible M according to the other, a vegetable is part of a plant that consists of the the edible part(s) of a plant, such as seeds and surrounding tissues. This the stems and stalk (celery), root includes fleshy fruits (such as blue- (carrot), tuber (potato), bulb (onion), berries, cantaloupe, poach, pumpkin, leaves (spinach, lettuce), flower (globe tomato) and dry fruits, where the artichoke), fruit (apple, cucumber, ripened ovary wall becomes papery, pumpkin, strawberries, tomato) or leathery, or woody as with cereal seeds (beans, peas). The latter grains, pulses (mature beans and definition includes fruits as a subset of peas) and nuts. vegetables. Definition of fruit and vegetables applicable in epidemiological studies, Fruit and vegetables Edible plant foods excluding -

The Mexican Cotton-Boll Weevil (Anthonomus Grandis Boh.)
Circular No. 14, Second Series. (Revision of No. 6.) United States Department of Agriculture, DIVISION OF ENTOMOLOGY. THE MEXICAN COTTON-BOLL WEEVIL. (Anthonomus grandis Bob.) SCOPE OF THE CIRCULAR. Circular No. 6 was published in April, 1895, and contained a brief report of the observations made up to that time, and the conclusions based on those observations, concerning the Mexican cotton boll weevil, an insect of Central American origin which, during 1894, attracted considera- ble attention in the cotton fields of south Texas. The investi- gation was continued during the summer, fall, and early winter of 1895, especially by Mr. Schwarz, who visited Texas in May and June and again from October to De- cember, and by Mr. Town send, who was FlG.l. Anthonomus grandis: a, adult beetle; b, pupa: larva—all enlarged. stationed in the State during the greater part of the summer. The writer went to Texas in December, and in companj- with Mr. Schwarz carefully studied the condition of affairs at that season and talked with many prominent cotton growers. The object of the present circular is to lay before cot- ton planters the results of this supplementary investigation. In order to make it complete in itself, such facts as are needed are repeated from Circular No. 6. GENERAL APPEARANCE AND METHOD OF WORK. This insect is a small, grayish weevil, of the shape and general appearance shown in fig. 1, «, and measuring a little less than a quar- ter of an inch in length. It is found in the cotton fields throughout the season, puncturing and laying its eggs in the squares and bolls.