Haloperidol Affects Coupling Between QT and RR Intervals in Guinea Pig Isolated Heart
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

List of Union Reference Dates A
Active substance name (INN) EU DLP BfArM / BAH DLP yearly PSUR 6-month-PSUR yearly PSUR bis DLP (List of Union PSUR Submission Reference Dates and Frequency (List of Union Frequency of Reference Dates and submission of Periodic Frequency of submission of Safety Update Reports, Periodic Safety Update 30 Nov. 2012) Reports, 30 Nov. -

)&F1y3x PHARMACEUTICAL APPENDIX to THE
)&f1y3X PHARMACEUTICAL APPENDIX TO THE HARMONIZED TARIFF SCHEDULE )&f1y3X PHARMACEUTICAL APPENDIX TO THE TARIFF SCHEDULE 3 Table 1. This table enumerates products described by International Non-proprietary Names (INN) which shall be entered free of duty under general note 13 to the tariff schedule. The Chemical Abstracts Service (CAS) registry numbers also set forth in this table are included to assist in the identification of the products concerned. For purposes of the tariff schedule, any references to a product enumerated in this table includes such product by whatever name known. Product CAS No. Product CAS No. ABAMECTIN 65195-55-3 ACTODIGIN 36983-69-4 ABANOQUIL 90402-40-7 ADAFENOXATE 82168-26-1 ABCIXIMAB 143653-53-6 ADAMEXINE 54785-02-3 ABECARNIL 111841-85-1 ADAPALENE 106685-40-9 ABITESARTAN 137882-98-5 ADAPROLOL 101479-70-3 ABLUKAST 96566-25-5 ADATANSERIN 127266-56-2 ABUNIDAZOLE 91017-58-2 ADEFOVIR 106941-25-7 ACADESINE 2627-69-2 ADELMIDROL 1675-66-7 ACAMPROSATE 77337-76-9 ADEMETIONINE 17176-17-9 ACAPRAZINE 55485-20-6 ADENOSINE PHOSPHATE 61-19-8 ACARBOSE 56180-94-0 ADIBENDAN 100510-33-6 ACEBROCHOL 514-50-1 ADICILLIN 525-94-0 ACEBURIC ACID 26976-72-7 ADIMOLOL 78459-19-5 ACEBUTOLOL 37517-30-9 ADINAZOLAM 37115-32-5 ACECAINIDE 32795-44-1 ADIPHENINE 64-95-9 ACECARBROMAL 77-66-7 ADIPIODONE 606-17-7 ACECLIDINE 827-61-2 ADITEREN 56066-19-4 ACECLOFENAC 89796-99-6 ADITOPRIM 56066-63-8 ACEDAPSONE 77-46-3 ADOSOPINE 88124-26-9 ACEDIASULFONE SODIUM 127-60-6 ADOZELESIN 110314-48-2 ACEDOBEN 556-08-1 ADRAFINIL 63547-13-7 ACEFLURANOL 80595-73-9 ADRENALONE -

Article (Published Version)
Article Characterization of drug‐induced transcriptional modules: towards drug repositioning and functional understanding ISKAR, Murat, et al. Reference ISKAR, Murat, et al. Characterization of drug‐induced transcriptional modules: towards drug repositioning and functional understanding. Molecular Systems Biology, 2013, vol. 9, no. 1, p. 662 DOI : 10.1038/msb.2013.20 PMID : 23632384 Available at: http://archive-ouverte.unige.ch/unige:153825 Disclaimer: layout of this document may differ from the published version. 1 / 1 Molecular Systems Biology 9; Article number 662; doi:10.1038/msb.2013.20 Citation: Molecular Systems Biology 9:662 www.molecularsystemsbiology.com Characterization of drug-induced transcriptional modules: towards drug repositioning and functional understanding Murat Iskar1, Georg Zeller1, Peter Blattmann2,3, Monica Campillos4,5, Michael Kuhn6, Katarzyna H Kaminska1,9, Heiko Runz3,7, Anne-Claude Gavin1, Rainer Pepperkok2,3, Vera van Noort1 and Peer Bork1,8,* 1 Structural and Computational Biology Unit, European Molecular Biology Laboratory (EMBL), Heidelberg, Germany, 2 Cell Biology/Biophysics Unit, EMBL, Heidelberg, Germany, 3 Molecular Medicine Partnership Unit (MMPU), EMBL, University of Heidelberg, Heidelberg, Germany, 4 Institute for Bioinformatics and Systems Biology, Helmholtz Center Munich–German Research Center for Environmental Health (GmbH), Neuherberg, Germany, 5 German Center for Diabetes Research (DZD), Neuherberg, Germany, 6 Biotechnology Center, TU Dresden, Dresden, Germany, 7 Institute of Human Genetics, University of Heidelberg, Heidelberg, Germany and 8 Max-Delbru¨ck-Centre for Molecular Medicine, Berlin, Germany 9Present address: International Institute of Molecular and Cell Biology in Warsaw, ul. Ks. Trojdena 4, 02-109 Warsaw, Poland * Corresponding author. Structural and Computational Biology Unit, European Molecular Biology Laboratory (EMBL), Meyerhofstrasse 1, Heidelberg, Germany. -

Etats Rapides
List of European Pharmacopoeia Reference Standards Effective from 2015/12/24 Order Reference Standard Batch n° Quantity Sale Information Monograph Leaflet Storage Price Code per vial Unit Y0001756 Exemestane for system suitability 1 10 mg 1 2766 Yes +5°C ± 3°C 79 ! Y0001561 Abacavir sulfate 1 20 mg 1 2589 Yes +5°C ± 3°C 79 ! Y0001552 Abacavir for peak identification 1 10 mg 1 2589 Yes +5°C ± 3°C 79 ! Y0001551 Abacavir for system suitability 1 10 mg 1 2589 Yes +5°C ± 3°C 79 ! Y0000055 Acamprosate calcium - reference spectrum 1 n/a 1 1585 79 ! Y0000116 Acamprosate impurity A 1 50 mg 1 3-aminopropane-1-sulphonic acid 1585 Yes +5°C ± 3°C 79 ! Y0000500 Acarbose 3 100 mg 1 See leaflet ; Batch 2 is valid until 31 August 2015 2089 Yes +5°C ± 3°C 79 ! Y0000354 Acarbose for identification 1 10 mg 1 2089 Yes +5°C ± 3°C 79 ! Y0000427 Acarbose for peak identification 3 20 mg 1 Batch 2 is valid until 31 January 2015 2089 Yes +5°C ± 3°C 79 ! A0040000 Acebutolol hydrochloride 1 50 mg 1 0871 Yes +5°C ± 3°C 79 ! Y0000359 Acebutolol impurity B 2 10 mg 1 -[3-acetyl-4-[(2RS)-2-hydroxy-3-[(1-methylethyl)amino] propoxy]phenyl] 0871 Yes +5°C ± 3°C 79 ! acetamide (diacetolol) Y0000127 Acebutolol impurity C 1 20 mg 1 N-(3-acetyl-4-hydroxyphenyl)butanamide 0871 Yes +5°C ± 3°C 79 ! Y0000128 Acebutolol impurity I 2 0.004 mg 1 N-[3-acetyl-4-[(2RS)-3-(ethylamino)-2-hydroxypropoxy]phenyl] 0871 Yes +5°C ± 3°C 79 ! butanamide Y0000056 Aceclofenac - reference spectrum 1 n/a 1 1281 79 ! Y0000085 Aceclofenac impurity F 2 15 mg 1 benzyl[[[2-[(2,6-dichlorophenyl)amino]phenyl]acetyl]oxy]acetate -

Stems for Nonproprietary Drug Names
USAN STEM LIST STEM DEFINITION EXAMPLES -abine (see -arabine, -citabine) -ac anti-inflammatory agents (acetic acid derivatives) bromfenac dexpemedolac -acetam (see -racetam) -adol or analgesics (mixed opiate receptor agonists/ tazadolene -adol- antagonists) spiradolene levonantradol -adox antibacterials (quinoline dioxide derivatives) carbadox -afenone antiarrhythmics (propafenone derivatives) alprafenone diprafenonex -afil PDE5 inhibitors tadalafil -aj- antiarrhythmics (ajmaline derivatives) lorajmine -aldrate antacid aluminum salts magaldrate -algron alpha1 - and alpha2 - adrenoreceptor agonists dabuzalgron -alol combined alpha and beta blockers labetalol medroxalol -amidis antimyloidotics tafamidis -amivir (see -vir) -ampa ionotropic non-NMDA glutamate receptors (AMPA and/or KA receptors) subgroup: -ampanel antagonists becampanel -ampator modulators forampator -anib angiogenesis inhibitors pegaptanib cediranib 1 subgroup: -siranib siRNA bevasiranib -andr- androgens nandrolone -anserin serotonin 5-HT2 receptor antagonists altanserin tropanserin adatanserin -antel anthelmintics (undefined group) carbantel subgroup: -quantel 2-deoxoparaherquamide A derivatives derquantel -antrone antineoplastics; anthraquinone derivatives pixantrone -apsel P-selectin antagonists torapsel -arabine antineoplastics (arabinofuranosyl derivatives) fazarabine fludarabine aril-, -aril, -aril- antiviral (arildone derivatives) pleconaril arildone fosarilate -arit antirheumatics (lobenzarit type) lobenzarit clobuzarit -arol anticoagulants (dicumarol type) dicumarol -

Achat Viagra Puissant ### Ablation Prostate Et Viagra >>> Kvadridze.Github.Io
Viagra est indiquée pour le traitement de la dysfonction érectile masculine. >>> ORDER NOW <<< Achat viagra puissant Tags: ordonnance de viagra combien de temps dur viagra comment peut on se procurer du viagra efficacité viagra générique un site fiable pour acheter du viagra le viagra sur ordonnance danger dacheter du viagra sur internet prix cachet viagra combien coute le viagra au maroc le rôle de viagra peut acheter du viagra sans ordonnance quesque cest viagra composition de viagra acheter viagra paris sans ordonnance comment se procurer viagra sans ordonnance comment reconnaitre le faux viagra quand faut il prendre viagra comment acheter du viagra au quebec lancement du viagra quest ce que cest le viagra viagra pfizer mode demploi les conséquences du viagra edex plus viagra comment prendre de viagra forum viagra pas cher commande de viagra en ligne meme effet que viagra prendre la moitié dun viagra experience viagra femme A vaginal ring can slip out of the vagina. In the event of overdosage, general symptomatic and achat viagra puissant measures are indicated as required Resistance to azithromycin may be inherent or acquired. Comments: Take one pill a day. Ca 20 minuter senare börjar mina kollegor droppa in i ordningen: Susanne, Britt, Emma, Nina, Gunnel, Eva och sist Barbro. Epidemiologic investigations of this outbreak demonstrated that individuals in close contact with the index case or with exposure to poultry were at risk of being infected. 25 mg pour femme viagra cherche 2 weeks or until I "felt" like I could go lower. It might take a while to adapt to it, since it will lower your blood pressure. -

Cardiac Sigma Receptors – an Update
Physiol. Res. 67 (Suppl. 4): S561-S576, 2018 https://doi.org/10.33549/physiolres.934052 REVIEW Cardiac Sigma Receptors – An Update T. STRACINA1, M. NOVAKOVA1 1Department of Physiology, Faculty of Medicine, Masaryk University, Brno, Czech Republic Received March 25, 2018 Accepted September 12, 2018 Summary (Martin et al. 1976). The authors believed that sigma More than four decades passed since sigma receptors were first receptor represents an opioid receptor subtype, which mentioned. Since then, existence of at least two receptor mediates psychomimetic and stimulatory behavioral subtypes and their tissue distributions have been proposed. effects of N-allylnormetazocine (SKF-10047) in chronic Nowadays, it is clear, that sigma receptors are unique ubiquitous spinal dog. Subsequent binding studies in guinea pig and proteins with pluripotent function, which can interact with so rat showed that binding profile of sigma receptor differs many different classes of proteins. As the endoplasmic resident from any other known subtype of opioid receptor as well proteins, they work as molecular chaperones – accompany as other receptor classes (Su 1982, Tam 1983). Therefore, various proteins during their folding, ensure trafficking of the the sigma receptor was defined as novel receptor type maturated proteins between cellular organelles and regulate their (Su 1982). functions. In the heart, sigma receptor type 1 is more dominant. Cardiac sigma 1 receptors regulate response to endoplasmic Two subtypes of sigma receptor reticulum stress, modulates calcium signaling in cardiomyocyte Further research led to differentiation among at and can affect function of voltage-gated ion channels. They least two subtypes of sigma receptors. Based on their contributed in pathophysiology of cardiac hypertrophy, heart diverse ligand selectivity and stereospecificity, association failure and many other cardiovascular disorders. -

Pharmaceutical Appendix to the Tariff Schedule 2
Harmonized Tariff Schedule of the United States (2007) (Rev. 2) Annotated for Statistical Reporting Purposes PHARMACEUTICAL APPENDIX TO THE HARMONIZED TARIFF SCHEDULE Harmonized Tariff Schedule of the United States (2007) (Rev. 2) Annotated for Statistical Reporting Purposes PHARMACEUTICAL APPENDIX TO THE TARIFF SCHEDULE 2 Table 1. This table enumerates products described by International Non-proprietary Names (INN) which shall be entered free of duty under general note 13 to the tariff schedule. The Chemical Abstracts Service (CAS) registry numbers also set forth in this table are included to assist in the identification of the products concerned. For purposes of the tariff schedule, any references to a product enumerated in this table includes such product by whatever name known. ABACAVIR 136470-78-5 ACIDUM LIDADRONICUM 63132-38-7 ABAFUNGIN 129639-79-8 ACIDUM SALCAPROZICUM 183990-46-7 ABAMECTIN 65195-55-3 ACIDUM SALCLOBUZICUM 387825-03-8 ABANOQUIL 90402-40-7 ACIFRAN 72420-38-3 ABAPERIDONUM 183849-43-6 ACIPIMOX 51037-30-0 ABARELIX 183552-38-7 ACITAZANOLAST 114607-46-4 ABATACEPTUM 332348-12-6 ACITEMATE 101197-99-3 ABCIXIMAB 143653-53-6 ACITRETIN 55079-83-9 ABECARNIL 111841-85-1 ACIVICIN 42228-92-2 ABETIMUSUM 167362-48-3 ACLANTATE 39633-62-0 ABIRATERONE 154229-19-3 ACLARUBICIN 57576-44-0 ABITESARTAN 137882-98-5 ACLATONIUM NAPADISILATE 55077-30-0 ABLUKAST 96566-25-5 ACODAZOLE 79152-85-5 ABRINEURINUM 178535-93-8 ACOLBIFENUM 182167-02-8 ABUNIDAZOLE 91017-58-2 ACONIAZIDE 13410-86-1 ACADESINE 2627-69-2 ACOTIAMIDUM 185106-16-5 ACAMPROSATE 77337-76-9 -
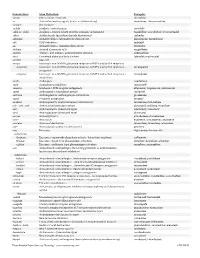
Downloaded from Survive Nursing | Survivenursing.Com V20110426
Generic Stem Stem Definition Examples -abine (see -arabine, -citabine) decitabine -ac Anti-inflammatory agents (acetic acid derivatives) bromfenac; dexpemedolac -acetam See -racetam -actide Synthetic corticotropins seractide -adol or -aldol- Analgesics (mixed opiate receptor agonists/ antagonists) tazadolene; spiradolene; levonantradol -adox Antibacterials (quinoline dioxide derivatives) carbadox -afenone Antiarrhythmics (propafenone derivatives) alprafenone; diprafenone -afil PDE5 inhibitors tadalafil -aj- Antiarrhythmics (ajmaline derivatives) lorajmine -aldrate Antacid aluminum salts magaldrate -algron Alpha1 - and alpha2 - adrenoreceptor agonists dabuzalgron -alol Combined alpha and beta blockers labetalol; medroxalol -amivir (see -vir) -ampa Ionotropic non-NMDA glutamate receptors (AMPA and/or KA receptors) -ampanel Ionotropic non-NMDA glutamate receptors (AMPA and/or KA receptors) ; becampanel antagonists -ampator Ionotropic non-NMDA glutamate receptors (AMPA and/or KA receptors) ; forampator modulators -andr- Androgens nandrolone -anib Angiogenesis inhibitors semaxanib -anserin Serotonin 5-HT2 receptor antagonists altanserin; tropanserin; adatanserin -antel Anthelmintics (undefined group) carbantel -antrone Antineoplastics; anthraquinone derivatives pixantrone -apsel P-selectin antagonists torapsel -arabine Antineoplastics (arabinofuranosyl derivatives) fazarabine; fludarabine aril-, -aril, -aril- Antiviral (arildone derivatives) pleconaril; arildone; fosarilate -arit Antirheumatics (lobenzarit type) lobenzarit; clobuzarit -arol -

Marrakesh Agreement Establishing the World Trade Organization
No. 31874 Multilateral Marrakesh Agreement establishing the World Trade Organ ization (with final act, annexes and protocol). Concluded at Marrakesh on 15 April 1994 Authentic texts: English, French and Spanish. Registered by the Director-General of the World Trade Organization, acting on behalf of the Parties, on 1 June 1995. Multilat ral Accord de Marrakech instituant l©Organisation mondiale du commerce (avec acte final, annexes et protocole). Conclu Marrakech le 15 avril 1994 Textes authentiques : anglais, français et espagnol. Enregistré par le Directeur général de l'Organisation mondiale du com merce, agissant au nom des Parties, le 1er juin 1995. Vol. 1867, 1-31874 4_________United Nations — Treaty Series • Nations Unies — Recueil des Traités 1995 Table of contents Table des matières Indice [Volume 1867] FINAL ACT EMBODYING THE RESULTS OF THE URUGUAY ROUND OF MULTILATERAL TRADE NEGOTIATIONS ACTE FINAL REPRENANT LES RESULTATS DES NEGOCIATIONS COMMERCIALES MULTILATERALES DU CYCLE D©URUGUAY ACTA FINAL EN QUE SE INCORPOR N LOS RESULTADOS DE LA RONDA URUGUAY DE NEGOCIACIONES COMERCIALES MULTILATERALES SIGNATURES - SIGNATURES - FIRMAS MINISTERIAL DECISIONS, DECLARATIONS AND UNDERSTANDING DECISIONS, DECLARATIONS ET MEMORANDUM D©ACCORD MINISTERIELS DECISIONES, DECLARACIONES Y ENTEND MIENTO MINISTERIALES MARRAKESH AGREEMENT ESTABLISHING THE WORLD TRADE ORGANIZATION ACCORD DE MARRAKECH INSTITUANT L©ORGANISATION MONDIALE DU COMMERCE ACUERDO DE MARRAKECH POR EL QUE SE ESTABLECE LA ORGANIZACI N MUND1AL DEL COMERCIO ANNEX 1 ANNEXE 1 ANEXO 1 ANNEX -

Customs Tariff - Schedule
CUSTOMS TARIFF - SCHEDULE 99 - i Chapter 99 SPECIAL CLASSIFICATION PROVISIONS - COMMERCIAL Notes. 1. The provisions of this Chapter are not subject to the rule of specificity in General Interpretative Rule 3 (a). 2. Goods which may be classified under the provisions of Chapter 99, if also eligible for classification under the provisions of Chapter 98, shall be classified in Chapter 98. 3. Goods may be classified under a tariff item in this Chapter and be entitled to the Most-Favoured-Nation Tariff or a preferential tariff rate of customs duty under this Chapter that applies to those goods according to the tariff treatment applicable to their country of origin only after classification under a tariff item in Chapters 1 to 97 has been determined and the conditions of any Chapter 99 provision and any applicable regulations or orders in relation thereto have been met. 4. The words and expressions used in this Chapter have the same meaning as in Chapters 1 to 97. Issued January 1, 2019 99 - 1 CUSTOMS TARIFF - SCHEDULE Tariff Unit of MFN Applicable SS Description of Goods Item Meas. Tariff Preferential Tariffs 9901.00.00 Articles and materials for use in the manufacture or repair of the Free CCCT, LDCT, GPT, UST, following to be employed in commercial fishing or the commercial MT, MUST, CIAT, CT, harvesting of marine plants: CRT, IT, NT, SLT, PT, COLT, JT, PAT, HNT, Artificial bait; KRT, CEUT, UAT, CPTPT: Free Carapace measures; Cordage, fishing lines (including marlines), rope and twine, of a circumference not exceeding 38 mm; Devices for keeping nets open; Fish hooks; Fishing nets and netting; Jiggers; Line floats; Lobster traps; Lures; Marker buoys of any material excluding wood; Net floats; Scallop drag nets; Spat collectors and collector holders; Swivels. -

(12) United States Patent (10) Patent No.: US 6,197,764 B1 Bradley Et Al
USOO6197764B1 (12) United States Patent (10) Patent No.: US 6,197,764 B1 Bradley et al. (45) Date of Patent: *Mar. 6, 2001 (54) CLOZAPINE COMPOSITIONS AND USES FOREIGN PATENT DOCUMENTS THEREOF 0599 576A1 1/1994 (EP). (75) Inventors: Matthews O. Bradley, Laytonsville, 693498 1/1996 (EP). MD (US); Victor E. Shashoua, 61204136 11/1984 (JP). Belmont, MA (US); Charles S. 06-072868 3/1994 (JP). Swindell, Merion; Nigel L. Webb, 6072868 3/1994 (JP). Bryn Mawr, both of PA (US) 7082146 3/1996 (JP). 8151334 6/1996 (JP). (73) Assignee: Protarga, Inc., Conshohocken, PA (US) 9030963 2/1997 (JP). (*) Notice: Subject to any disclaimer, the term of this WO 89/07938 9/1989 (WO). patent is extended or adjusted under 35 WO 96/04001 2/1996 (WO). U.S.C. 154(b) by 0 days. WO 96/22303 7/1996 (WO). WO 96/27380 9/1996 (WO). This patent is Subject to a terminal dis WO98/17325 4/1998 (WO). claimer. OTHER PUBLICATIONS (21) Appl. No.: 08/978,541 (22) Filed: Nov. 26, 1997 Bourat, et al., "Long Chain Esters of Pipotiazine as Lon g-Acting Psychotropic Pro-Drug, Med. Chem. Proc. Int. (51) Int. Cl. .............................................. A61K 31/00 Symp. 5th (1976) pp. 105-114. (52) U.S. Cl. ........................... 514/218; 514/219; 514/220 Lohr, et al., “Neuroleptic-Induced Movement Disorders . (58) Field of Search ..................................... 514/218, 219, ..", Psychiatry, vol. 3, (1989). 514/220 Makino, et al., Chemical Abstracts, vol. 106, No. 12, (56) References Cited (90.177x) issued Mar. 23, 1987, “Pharmaceuticals Permeable to Blood-Brain Barrier'.