Intramedullary Spinal Cord Tumors: Molecular Insights and Surgical Innovation
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Vetp-42-06-Brief Communications
Veterinary Pathology Online http://vet.sagepub.com/ Primitive Neuroectodermal Tumor in the Spinal Cord of a Brahman Crossbred Calf A. Berrocal, D. L. Montgomery, J. T. Mackie and R. W. Storts Vet Pathol 2005 42: 834 DOI: 10.1354/vp.42-6-834 The online version of this article can be found at: http://vet.sagepub.com/content/42/6/834 Published by: http://www.sagepublications.com On behalf of: American College of Veterinary Pathologists, European College of Veterinary Pathologists, & the Japanese College of Veterinary Pathologists. Additional services and information for Veterinary Pathology Online can be found at: Email Alerts: http://vet.sagepub.com/cgi/alerts Subscriptions: http://vet.sagepub.com/subscriptions Reprints: http://www.sagepub.com/journalsReprints.nav Permissions: http://www.sagepub.com/journalsPermissions.nav Downloaded from vet.sagepub.com by guest on December 25, 2010 834 Brief Communications and Case Reports Vet Pathol 42:6, 2005 Vet Pathol 42:834–836 (2005) Primitive Neuroectodermal Tumor in the Spinal Cord of a Brahman Crossbred Calf A. BERROCAL,D.L.MONTGOMERY,J.T.MACKIE, AND R. W. STORTS Abstract. A variety of embryonal tumors of the central nervous system, typically malignant and occurring in young individuals, are recognized in humans and animals. This report describes an invasive subdural but predominantly extramedullary primitive neuroectodermal tumor developing at the lumbosacral junction in a 6-month-old Brahman crossbred calf. The tumor was composed of spindloid embryonal cells organized in interlacing fascicles. The cells had oval to elongate or round hyperchromic nuclei, single to double nucleoli, and scant discernible cytoplasm. Immunohistochemical staining for neuron-specific enolase, synaptophysin, and S-100 protein and formation of pseudorosettes suggested neuronal and possibly ependymal differentiation. -

Intraventricular Neuroepithelial Tumors: Surgical Outcome, Technical Considerations and Review of Literature A
Aftahy et al. BMC Cancer (2020) 20:1060 https://doi.org/10.1186/s12885-020-07570-1 RESEARCH ARTICLE Open Access Intraventricular neuroepithelial tumors: surgical outcome, technical considerations and review of literature A. Kaywan Aftahy1* , Melanie Barz1, Philipp Krauss1, Friederike Liesche2, Benedikt Wiestler3, Stephanie E. Combs4,5,6, Christoph Straube4, Bernhard Meyer1 and Jens Gempt1 Abstract Background: Intraventricular neuroepithelial tumors (IVT) are rare lesions and comprise different pathological entities such as ependymomas, subependymomas and central neurocytomas. The treatment of choice is neurosurgical resection, which can be challenging due to their intraventricular location. Different surgical approaches to the ventricles are described. Here we report a large series of IVTs, its postoperative outcome at a single tertiary center and discuss suitable surgical approaches. Methods: We performed a retrospective chart review at a single tertiary neurosurgical center between 03/2009–05/ 2019. We included patients that underwent resection of an IVT emphasizing on surgical approach, extent of resection, clinical outcome and postoperative complications. Results: Forty five IVTs were resected from 03/2009 to 05/2019, 13 ependymomas, 21 subependymomas, 10 central neurocytomas and one glioependymal cyst. Median age was 52,5 years with 55.6% (25) male and 44.4% (20) female patients. Gross total resection was achieved in 93.3% (42/45). 84.6% (11/13) of ependymomas, 100% (12/21) of subependymomas, 90% (9/10) of central neurocytomas and one glioependymal cyst were completely removed. Postoperative rate of new neurological deficits was 26.6% (12/45). Postoperative new permanent cranial nerve deficits occurred in one case with 4th ventricle subependymoma and one in 4th ventricle ependymoma. -

Split Spinal Cord Malformations in Children
Split spinal cord malformations in children Yusuf Ersahin, M.D., Saffet Mutluer, M.D., Sevgül Kocaman, R.N., and Eren Demirtas, M.D. Division of Pediatric Neurosurgery, Department of Neurosurgery, and Department of Pathology, Ege University Faculty of Medicine, Izmir, Turkey The authors reviewed and analyzed information on 74 patients with split spinal cord malformations (SSCMs) treated between January 1, 1980 and December 31, 1996 at their institution with the aim of defining and classifying the malformations according to the method of Pang, et al. Computerized tomography myelography was superior to other radiological tools in defining the type of SSCM. There were 46 girls (62%) and 28 boys (38%) ranging in age from less than 1 day to 12 years (mean 33.08 months). The mean age (43.2 months) of the patients who exhibited neurological deficits and orthopedic deformities was significantly older than those (8.2 months) without deficits (p = 0.003). Fifty-two patients had a single Type I and 18 patients a single Type II SSCM; four patients had composite SSCMs. Sixty-two patients had at least one associated spinal lesion that could lead to spinal cord tethering. After surgery, the majority of the patients remained stable and clinical improvement was observed in 18 patients. The classification of SSCMs proposed by Pang, et al., will eliminate the current chaos in terminology. In all SSCMs, either a rigid or a fibrous septum was found to transfix the spinal cord. There was at least one unrelated lesion that caused tethering of the spinal cord in 85% of the patients. -

Charts Chart 1: Benign and Borderline Intracranial and CNS Tumors Chart
Charts Chart 1: Benign and Borderline Intracranial and CNS Tumors Chart Glial Tumor Neuronal and Neuronal‐ Ependymomas glial Neoplasms Subependymoma Subependymal Giant (9383/1) Cell Astrocytoma(9384/1) Myyppxopapillar y Desmoplastic Infantile Ependymoma Astrocytoma (9412/1) (9394/1) Chart 1: Benign and Borderline Intracranial and CNS Tumors Chart Glial Tumor Neuronal and Neuronal‐ Ependymomas glial Neoplasms Subependymoma Subependymal Giant (9383/1) Cell Astrocytoma(9384/1) Myyppxopapillar y Desmoplastic Infantile Ependymoma Astrocytoma (9412/1) (9394/1) Use this chart to code histology. The tree is arranged Chart Instructions: Neuroepithelial in descending order. Each branch is a histology group, starting at the top (9503) with the least specific terms and descending into more specific terms. Ependymal Embryonal Pineal Choro id plexus Neuronal and mixed Neuroblastic Glial Oligodendroglial tumors tumors tumors tumors neuronal-glial tumors tumors tumors tumors Pineoblastoma Ependymoma, Choroid plexus Olfactory neuroblastoma Oligodendroglioma NOS (9391) (9362) carcinoma Ganglioglioma, anaplastic (9522) NOS (9450) Oligodendroglioma (9390) (9505 Olfactory neurocytoma Ganglioglioma, malignant (()9521) anaplastic (()9451) Anasplastic ependymoma (9505) Olfactory neuroepithlioma Oligodendroblastoma (9392) (9523) (9460) Papillary ependymoma (9393) Glioma, NOS (9380) Supratentorial primitive Atypical EdEpendymo bltblastoma MdllMedulloep ithliithelioma Medulloblastoma neuroectodermal tumor tetratoid/rhabdoid (9392) (9501) (9470) (PNET) (9473) tumor -

Central Nervous System Tumors General ~1% of Tumors in Adults, but ~25% of Malignancies in Children (Only 2Nd to Leukemia)
Last updated: 3/4/2021 Prepared by Kurt Schaberg Central Nervous System Tumors General ~1% of tumors in adults, but ~25% of malignancies in children (only 2nd to leukemia). Significant increase in incidence in primary brain tumors in elderly. Metastases to the brain far outnumber primary CNS tumors→ multiple cerebral tumors. One can develop a very good DDX by just location, age, and imaging. Differential Diagnosis by clinical information: Location Pediatric/Young Adult Older Adult Cerebral/ Ganglioglioma, DNET, PXA, Glioblastoma Multiforme (GBM) Supratentorial Ependymoma, AT/RT Infiltrating Astrocytoma (grades II-III), CNS Embryonal Neoplasms Oligodendroglioma, Metastases, Lymphoma, Infection Cerebellar/ PA, Medulloblastoma, Ependymoma, Metastases, Hemangioblastoma, Infratentorial/ Choroid plexus papilloma, AT/RT Choroid plexus papilloma, Subependymoma Fourth ventricle Brainstem PA, DMG Astrocytoma, Glioblastoma, DMG, Metastases Spinal cord Ependymoma, PA, DMG, MPE, Drop Ependymoma, Astrocytoma, DMG, MPE (filum), (intramedullary) metastases Paraganglioma (filum), Spinal cord Meningioma, Schwannoma, Schwannoma, Meningioma, (extramedullary) Metastases, Melanocytoma/melanoma Melanocytoma/melanoma, MPNST Spinal cord Bone tumor, Meningioma, Abscess, Herniated disk, Lymphoma, Abscess, (extradural) Vascular malformation, Metastases, Extra-axial/Dural/ Leukemia/lymphoma, Ewing Sarcoma, Meningioma, SFT, Metastases, Lymphoma, Leptomeningeal Rhabdomyosarcoma, Disseminated medulloblastoma, DLGNT, Sellar/infundibular Pituitary adenoma, Pituitary adenoma, -

Cervical Myelopathy • Pragmatic Review
DIAGNOSIS AND INDICATIONS FOR SURGERY Cervical Spondylitic Myelopathy Timothy A. Garvey, MD Twin Cities Spine Center Minneapolis, MN Cervical Myelopathy • Pragmatic review • Differential diagnosis • History and physical exam • Reality check – clinical cases last few months • Every-day clinical decisions Cervical Myelopathy • Degenerative • Cervical spondylosis, stenosis, OPLL • Trauma • SCI, fracture • Tumor • Neoplasms • Autoimmune • MS, ALS, SLE, etc. • Congenital • Syrinx, Chiari, Down’s Cervical Myelopathy • Infection • Viral: HIV, polio, herpes • Bacterial: Syphilis, epidural abscess • Arthritis • RA, SLE. Sjogren • Vascular • Trauma, AVM, Anklosing spondylitis • Metabolic • Vitamin, liver, gastric bypass, Hepatitis C • Toxins • Methalene blue, anesthetics • Misc. • Radiation, baro trauma, electrical injury Myelopathy • Generic spinal cord dysfunction • Classification systems • Nurick, Ranawat, JOA • Nice review • CSRS 2005, Chapter 15 – Lapsiwala & Trost Cervical Myelopathy • Symptoms • Gait disturbance Ataxia • Weakness, hand function • Sensory – numbness and tingling • Bladder - urgency Cervical Myelopathy • Signs • Reflexes – hyper-reflexia, clonus • Pathologic – Hoffman’s, Babinski • Motor – weakness • Sensory – variable • Ataxia – unsteady gait • Provocative – Lhrmitte’s Pathophysiology/CSM • Cord compression with distortion • Ischemia – anterior spinal flow • Axoplasmic flow diminution • Demyelinization of the white matter in both ascending and descending traits L.F. • 48 year-old female w/work injury • “cc” – left leg dysfunction – Mild urinary frequency – More weakness than pain • PE – Mild weakness, multiple groups – No UMN L.F. 08/06 L4-5 L2-3 L.F. 09/07 L.F. 1/07 L.F. 7/07 Asymptomatic MRI • 100 patients • Disc Protrusion – 20% of 45 – 54 year olds – 57% of > 64 year olds • Cord Impingement – 16% < 64 – 26% > 64 • Cord Compression – 7 of 100 Asymptomatic Degenerative Disk Disease and Spondylosis of the Cervical Spine: MR Imaging. -

Spinal Injury
SPINAL INJURY Presented by:- Bhagawati Ray DEFINITION Spinal cord injury (SCI) is damage to the spinal cord that results in a loss of function such as mobility or feeling. TYPES OF SPINAL CORD INJURY Complete Spinal Cord Injuries Complete paraplegia is described as permanent loss of motor and nerve function at T1 level or below, resulting in loss of sensation and movement in the legs, bowel, bladder, and sexual region. Arms and hands retain normal function. INCOMPLETE SPINAL CORD INJURIES Anterior cord syndrome Anterior cord syndrome, due to damage to the front portion of the spinal cord or reduction in the blood supply from the anterior spinal artery, can be caused by fractures or dislocations of vertebrae or herniated disks. CENTRAL CORD SYNDROME Central cord syndrome, almost always resulting from damage to the cervical spinal cord, is characterized by weakness in the arms with relative sparing of the legs, and spared sensation in regions served by the sacral segments. POSTERIOR CORD SYNDROME Posterior cord syndrome, in which just the dorsal columns of the spinal cord are affected, is usually seen in cases of chronic myelopathy but can also occur with infarction of the posterior spinal artery. BROWN-SEQUARD SYNDROME Brown-Sequard syndrome occurs when the spinal cord is injured on one side much more than the other. It is rare for the spinal cord to be truly hemisected (severed on one side), but partial lesions due to penetrating wounds (such as gunshot or knife wounds) or fractured vertebrae or tumors are common. CAUDA EQUINASYNDROME Cauda equina syndrome (CES) is a condition that occurs when the bundle of nerves below the end of the spinal cord known as the cauda equina is damaged. -

Radiation Oncology Guidelines
National Imaging Associates, Inc.* 2021 Magellan Clinical Guidelines For Medical Necessity Review RADIATION ONCOLOGY GUIDELINES Effective January 1, 2021 – December 31, 2021 *National Imaging Associates, Inc. (NIA) is a subsidiary of Magellan Healthcare, Inc. Copyright © 2019-2020 National Imaging Associates, Inc., All Rights Reserved Guidelines for Clinical Review Determination Preamble Magellan is committed to the philosophy of supporting safe and effective treatment for patients. The medical necessity criteria that follow are guidelines for the provision of diagnostic imaging. These criteria are designed to guide both providers and reviewers to the most appropriate diagnostic tests based on a patient’s unique circumstances. In all cases, clinical judgment consistent with the standards of good medical practice will be used when applying the guidelines. Determinations are made based on both the guideline and clinical information provided at the time of the request. It is expected that medical necessity decisions may change as new evidence-based information is provided or based on unique aspects of the patient’s condition. The treating clinician has final authority and responsibility for treatment decisions regarding the care of the patient. 2021 Magellan Clinical Guidelines-Radiation Oncology 2 Guideline Development Process These medical necessity criteria were developed by Magellan Healthcare for the purpose of making clinical review determinations for requests for therapies and diagnostic procedures. The developers of the criteria sets included representatives from the disciplines of radiology, internal medicine, nursing, cardiology, and other specialty groups. Magellan’s guidelines are reviewed yearly and modified when necessary following a literature search of pertinent and established clinical guidelines and accepted diagnostic imaging practices. -

Spinal Meningiomas: a Review
nal of S ur pi o n J e Galgano et al., J Spine 2014, 3:1 Journal of Spine DOI: 10.4172/2165-7939.1000157 ISSN: 2165-7939 Review Rrticle Open Access Spinal Meningiomas: A Review Michael A Galgano, Timothy Beutler, Aaron Brooking and Eric M Deshaies* Department of Neurosurgery, SUNY Upstate Medical University, Syracuse, NY, USA Abstract Meningiomas of the spinal axis have been identified from C1 to as distal as the sacrum. Their clinical presentation varies greatly based on their location. Meningiomas situated in the atlanto-axial region may present similarly to some meningiomas of the craniocervical junction, while some of the more distal spinal axis meningiomas are discovered as a result of chronic back pain. Surgical resection remains the mainstay of treatment, although advancements in radiosurgery have led to increased utilization as a primary or adjuvant therapy. Angiography also plays a critical role in surgical planning and may be utilized for preoperative embolization of hypervascular meningiomas. Keywords: Meningioma; Neurosurgery; Radiosurgery; Angiography favorable and biological outcome [11]. They also found that either a lack of estrogen and progesterone receptors, or the presence of estrogen Introduction receptors in meningiomas, correlated with a more aggressive clinical Spinal meningiomas are tumors originating from arachnoid cap behavior, progression, and recurrence. Hsu and Hedley Whyte also cells most commonly situated in the intradural extramedullary region found that the presence of progesterone receptors, even in a small [1,2]. They represent a high proportion of all spinal cord tumors. Spinal subgroup of tumor cells, indicated a more favorable prognostic value meningiomas tend to predominate in the thoracic region, although for meningiomas [12]. -

Syringomyelia in Cervical Spondylosis: a Rare Sequel H
THIEME Editorial 1 Editorial Syringomyelia in Cervical Spondylosis: A Rare Sequel H. S. Bhatoe1 1 Department of Neurosciences, Max Super Specialty Hospital, Patparganj, New Delhi, India Indian J Neurosurg 2016;5:1–2. Neurological involvement in cervical spondylosis usually the buckled hypertrophic ligament flavum compresses the implies radiculopathy or myelopathy. Cervical spondylotic cord. Ischemia due to compromise of microcirculation and myelopathy is the commonest cause of myelopathy in the venous congestion, leading to focal demyelination.3 geriatric age group,1 and often an accompaniment in adult Syringomyelia is an extremely rare sequel of chronic cervical patients manifesting central cord syndrome and spinal cord cord compression due to spondylotic process, and manifests as injury without radiographic abnormality. Myelopathy is the accelerated myelopathy (►Fig. 1). Pathogenesis of result of three factors that often overlap: mechanical factors, syringomyelia is uncertain. Al-Mefty et al4 postulated dynamic-repeated microtrauma, and ischemia of spinal cord occurrence of myelomalacia due to chronic compression of microcirculation.2 Age-related mechanical changes include the cord, followed by phagocytosis, leading to a formation of hypertrophy of the ligamentum flavum, formation of the cavity that extends further. However, Kimura et al5 osteophytic bars, degenerative disc prolapse, all of them disagreed with this hypothesis, and postulated that following contributing to a narrowing of the spinal canal. Degenerative compression of the cord, there is slosh effect cranially and kyphosis and subluxation often aggravates the existing caudally, leading to an extension of the syrinx. It is thus likely compressiononthespinalcord.Flexion–extension that focal cord cavitation due to compression and ischemia movements of the spinal cord places additional, dynamic occurs due to periventricular fluid egress into the cord, the stretch on the cord that is compressed. -

Cerebellar Anaplastic Astrocytoma in Adult Patients: 15 Consecutive Cases from a Single Institution and Literature Review
medRxiv preprint doi: https://doi.org/10.1101/2020.09.09.20188938; this version posted September 14, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in perpetuity. All rights reserved. No reuse allowed without permission. Cerebellar anaplastic astrocytoma in adult patients: 15 consecutive cases from a single institution and literature review Artem Belyaev1, Dmitry Usachev1, Marina Ryzhova1, Gleb Gulida1, Vasilisa Skvortsova2,3, Igor Pronin1, Grigory Kobiakov1. 1 Burdenko Neurosurgery Center, 4th Tverskaya-Yamskaya, 16, Moscow, 125047, Russia 2 Wellcome Trust Centre for Neuroimaging, University College London, London, United Kingdom; 3 Max Planck University College London Centre for Computational Psychiatry and Ageing Research, London, United Kingdom; Abstract: Adult cerebellar anaplastic astrocytomas (cAA) are rare entities and their clinical and genetic appearances are still ill defined. Previously, malignant gliomas of the cerebellum were combined and reviewed together (cAA and cerebellar glioblastomas (cGB), that could have possibly affected overall results. We present characteristics of 15 adult patients with cAA and compared them to a series of 45 patients with a supratentorial AA (sAA). The mean age at cAA diagnosis was 39.3 years (range 19-72). A history of neurofibromatosis type I was noted in 1 patient (6.7%). An IDH-1 mutation was identified in 6/15 cases and a methylated MGMT promoter in 5/15 cases. Patients in study and control groups were matched in age, sex and IDH- 1 mutation status. Patients in a study group tended to have a more frequent multifocal presentation at diagnosis (13% vs. -
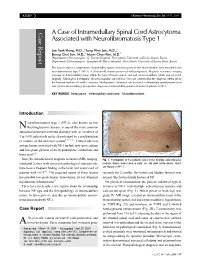
A Case of Intramedullary Spinal Cord Astrocytoma Associated with Neurofibromatosis Type 1
KISEP J Korean Neurosurg Soc 36 : 69-71, 2004 Case Report A Case of Intramedullary Spinal Cord Astrocytoma Associated with Neurofibromatosis Type 1 Jae Taek Hong, M.D.,1 Sang Won Lee, M.D.,1 Byung Chul Son, M.D.,1 Moon Chan Kim, M.D.2 Department of Neurosurgery,1 St. Vincent Hospital, The Catholic University of Korea, Suwon, Korea Department of Neurosurgery,2 Kangnam St. Mary's Hospital, The Catholic University of Korea, Seoul, Korea The authors report a symptomatic intramedullary spinal cord astrocytoma in the thoracolumbar area associated with neurofibromatosis type 1 (NF-1). A 38-year-old woman presented with paraparesis. Magnetic resonance imaging revealed an intramedullary lesion within the lower thoracic spinal cord and conus medullaris, which was removed surgically. Pathological investigation showed anaplastic astrocytoma. This case confirms that the diagnosis criteria set by the National Institute of Health Consensus Development Conference can be useful to differentiate ependymoma from astrocytoma when making a preoperative diagnosis of intramedullary spinal cord tumor in patients of NF-1. KEY WORDS : Astrocytoma·Intramedullary cord tumor·Neurofibromatosis. Introduction eurofibromatosis type 1 (NF-1), also known as von N Recklinghausen's disease, is one of the most common autosomal dominant inherited disorders with an incidence of 1 in 3,000 individuals and is characterized by a predisposition to tumors of the nervous system5,6,12,16). Central nervous system lesions associated with NF-1 include optic nerve glioma and low-grade gliomas of the hypothalamus, cerebellum and brain stem6,10). Since the introduction of magnetic resonance(MR) imaging, Fig. 1. Photograph of the patient's back shows multiple subcutaneous incidental lesions with uncertain pathological characteristic nodules (black arrow) and a cafe-au-lait spot (white arrow), which have been a frequent finding in the brain and spinal cord of are typical of NF-1.