L/Ba1ercanjiuscum
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Xhaie'ican%Mllsllm
XhAie'ican1ox4tate%Mllsllm PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK 24, N.Y. NUMBER 2 244 MAY I9, I 966 The Larvae of the Anthophoridae (Hymenoptera, Apoidea) Part 2. The Nomadinae BY JEROME G. ROZEN, JR.1 The present paper is the second of a series that treats the phylogeny and taxonomy of the larvae belonging to the bee family Anthophoridae. The first (Rozen, 1965a) dealt with the pollen-collecting tribes Eucerini and Centridini of the Anthophorinae. The present study encompasses the following tribes, all of which consist solely of cuckoo bees: Protepeolini, Epeolini, Nomadini, Ammobatini, Holcopasitini, Biastini, and Neolarrini. For reasons presented below, these tribes are believed to represent a monophyletic group, and consequently all are placed in the Nomadinae. It seems likely that the cleptoparasitic tribes Caenoprosopini, Ammoba- toidini, Townsendiellini, Epeoloidini, and Osirini are also members of the subfamily, although their larvae have not as yet been collected. Although the interrelationships of the numerous taxa within the Nomadinae need to be re-evaluated, the tribal concepts used by Michener (1944) are employed here. Adjustments in the classifications will certainly have to be made in the future, however, for Michener (1954) has already indicated, for example, that characters of the adults in the Osirini, the Epeolini, and the Nomadini intergrade. The affinities of the Nomadinae with the other subfamilies of the Antho- phoridae will be discussed in the last paper of the series. Because of char- 1 Curator, Department of Entomology, the American Museum of Natural History. 2 AMERICAN MUSEUM NOVITATES NO. -

Bees of the Genus Panurginus in Siberia, Far East of Russia, and Allied Areas (Hymenoptera: Andrenidae, Panurginae)
Zootaxa 3112: 1–35 (2011) ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ Article ZOOTAXA Copyright © 2011 · Magnolia Press ISSN 1175-5334 (online edition) Bees of the genus Panurginus in Siberia, Far East of Russia, and Allied Areas (Hymenoptera: Andrenidae, Panurginae) TATIANA G. ROMANKOVA1 & YULIA V. ASTAFUROVA2 1E-mail: [email protected] 2Zoological Institute, Russian Academy of Sciences, Universitetskaya Nab., 1, St. Petersburg, 199034, Russia. E-mail: [email protected] Table of contents Introduction . 2 Material and methods . 2 Genus Panurginus Nylander, 1848 . 2 Discussion . 3 Key for the genus Panurginus of Siberia, Far East of Russia and allied areas . 5 Annotated list of species . 7 1. Panurginus alticolus F. Morawitz . 7 2. Panurginus alpotanini sp. n. 9 3. Panurginus arsenievi sp. n. 15 4. Panurginus crawfordi Cockerell . 17 5. Panurginus herzi F. Morawitz . 19 6. Panurginus kozlovi sp. n. 20 7. Panurginus kropotkini sp. n. 22 8. Panurginus labiatus (Eversmann) . 23 9. Panurginus mikhno sp. n. 25 10. Panurginus muraviovi sp. n. 26 11. Panurginus niger Nylander . 27 12. Panurginus nigripes F. Morawitz . 29 13. Panurginus obruchevi sp. n. 30 14. Panurginus romani Aurivillius . 31 Dedication and acknowledgements . 33 References . 33 Abstract The genus Panurginus Nylander of the Asian part of Palearctic is revised for the first time. 14 species are listed for Siberia, the Far East of Russia, Kazakhstan, Mongolia, and Northern China. Lectotypes for P. alticolus F. Morawitz, 1876, P. herzi F. Morawitz, 1891 and P. labiatus Eversmann, 1852 are designated. Panurginus niger Nylander, 1848, the type species of the genus, was studied for the first time after it was described. -

Diversity and Resource Choice of Flower-Visiting Insects in Relation to Pollen Nutritional Quality and Land Use
Diversity and resource choice of flower-visiting insects in relation to pollen nutritional quality and land use Diversität und Ressourcennutzung Blüten besuchender Insekten in Abhängigkeit von Pollenqualität und Landnutzung Vom Fachbereich Biologie der Technischen Universität Darmstadt zur Erlangung des akademischen Grades eines Doctor rerum naturalium genehmigte Dissertation von Dipl. Biologin Christiane Natalie Weiner aus Köln Berichterstatter (1. Referent): Prof. Dr. Nico Blüthgen Mitberichterstatter (2. Referent): Prof. Dr. Andreas Jürgens Tag der Einreichung: 26.02.2016 Tag der mündlichen Prüfung: 29.04.2016 Darmstadt 2016 D17 2 Ehrenwörtliche Erklärung Ich erkläre hiermit ehrenwörtlich, dass ich die vorliegende Arbeit entsprechend den Regeln guter wissenschaftlicher Praxis selbständig und ohne unzulässige Hilfe Dritter angefertigt habe. Sämtliche aus fremden Quellen direkt oder indirekt übernommene Gedanken sowie sämtliche von Anderen direkt oder indirekt übernommene Daten, Techniken und Materialien sind als solche kenntlich gemacht. Die Arbeit wurde bisher keiner anderen Hochschule zu Prüfungszwecken eingereicht. Osterholz-Scharmbeck, den 24.02.2016 3 4 My doctoral thesis is based on the following manuscripts: Weiner, C.N., Werner, M., Linsenmair, K.-E., Blüthgen, N. (2011): Land-use intensity in grasslands: changes in biodiversity, species composition and specialization in flower-visitor networks. Basic and Applied Ecology 12 (4), 292-299. Weiner, C.N., Werner, M., Linsenmair, K.-E., Blüthgen, N. (2014): Land-use impacts on plant-pollinator networks: interaction strength and specialization predict pollinator declines. Ecology 95, 466–474. Weiner, C.N., Werner, M , Blüthgen, N. (in prep.): Land-use intensification triggers diversity loss in pollination networks: Regional distinctions between three different German bioregions Weiner, C.N., Hilpert, A., Werner, M., Linsenmair, K.-E., Blüthgen, N. -

The Bees of Sub-Saharan Africa
A-PDF Split DEMO : Purchase from www.A-PDF.com to remove the watermark Genus Nasutapis Michener (Fig. 36E) Nasutapis has a distinct projection medioventrally on the clypeus. This genus is monotypic (Nasutapis straussorum Michener) and endemic to KwaZulu-Natal, South Africa, and found in nests of Braunsapis facialis (Gerstaecker). 8.6.2. Subfamily Nomadinae In sub-Saharan Africa the Nomadinae comprises four tribes and six genera. They are all cleptoparasitic. Diagnostic features for the subfamily are difficult to define, but almost each tribe has a distinctive feature, except Ammobatoidini. 8.6.2.1. Tribe Nomadini Genus Nomada Scopoli (Fig. 37A) Nomadini has one genus in sub-Saharan Africa, namely Nomada. There are ten species, occurring mostly in North-East and southern Africa. 8.6.2.2. Tribe Epeolini Genus Epeolus Latreille (Fig. 37B) Epeolini has one genus in sub-Saharan Africa, namely Epeolus. There are 13 species that occur mostly on the east side of the continent, along its entire length. 8.6.2.3. Tribe Ammobatoidini Genus Ammobatoides Radoszkowski (Fig. 37C) Ammobatoidini has one genus in sub-Saharan Africa, and it is known only from the holotype of Ammobatoides braunsi Bischoff. It was collected in Willowmore, South Africa. It therefore goes without saying that it is extremely rare. 8.6.2.4. Tribe Ammobatini The Ammobatini has four sub-Saharan genera. They all comprise cleptoparasitic bees. Ammobates has its centre of diversity in the Palaearctic, as does Chiasmognathus, which occurs just north of the Afrotropical Region and intrudes into sub-Saharan Africa. Pasites is mostly Afrotropical and Sphecodopsis is endemic to southern Africa. -

Pollination of Cultivated Plants in the Tropics 111 Rrun.-Co Lcfcnow!Cdgmencle
ISSN 1010-1365 0 AGRICULTURAL Pollination of SERVICES cultivated plants BUL IN in the tropics 118 Food and Agriculture Organization of the United Nations FAO 6-lina AGRICULTUTZ4U. ionof SERNES cultivated plans in tetropics Edited by David W. Roubik Smithsonian Tropical Research Institute Balboa, Panama Food and Agriculture Organization of the United Nations F'Ø Rome, 1995 The designations employed and the presentation of material in this publication do not imply the expression of any opinion whatsoever on the part of the Food and Agriculture Organization of the United Nations concerning the legal status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. M-11 ISBN 92-5-103659-4 All rights reserved. No part of this publication may be reproduced, stored in a retrieval system, or transmitted in any form or by any means, electronic, mechanical, photocopying or otherwise, without the prior permission of the copyright owner. Applications for such permission, with a statement of the purpose and extent of the reproduction, should be addressed to the Director, Publications Division, Food and Agriculture Organization of the United Nations, Viale delle Terme di Caracalla, 00100 Rome, Italy. FAO 1995 PlELi. uion are ted PlauAr David W. Roubilli (edita Footli-anal ISgt-iieulture Organization of the Untled Nations Contributors Marco Accorti Makhdzir Mardan Istituto Sperimentale per la Zoologia Agraria Universiti Pertanian Malaysia Cascine del Ricci° Malaysian Bee Research Development Team 50125 Firenze, Italy 43400 Serdang, Selangor, Malaysia Stephen L. Buchmann John K. S. Mbaya United States Department of Agriculture National Beekeeping Station Carl Hayden Bee Research Center P. -

Apoidea: Andrenidae: Panurginae)
Contemporary distributions of Panurginus species and subspecies in Europe (Apoidea: Andrenidae: Panurginae) Sébastien Patiny Abstract The largest number of Old World Panurginus Nylander, 1848 species is distributed in the West- Palaearctic. The genus is absent in Africa and rather rare in the East-Palaearctic. Warncke (1972, 1987), who is the main author treating Palaearctic Panurginae in the last decades, subdivided Panurginus into a small number of species, including two principal taxa admitting for each a very large number of subspecies: Panurginus brullei (Lepeletier, 1841) and Panurginus montanus Giraud, 1861. Following recent works, the two latter are in fact complexes of closely related species. In the West-Palaearctic context, distributions of certain species implied in these complexes appear as very singular, distinct of mostly all other Panurginae ranges and highly interesting from a fundamental point of view. Based on a cartographic approach, the causes which influence (or have conditioned, in the past) the observed ranges of these species are discussed. Key words: Panurginae, biogeography, West-Palaearctic, speciation, glaciation. Introduction 1841) sensu lato are studied in the present paper. In the Old World Panurginae fauna, Panurginus The numerous particularities of their distribu- Nylander, 1848 is one of the only two genera tions are characterized and discussed here. The (with Melitturga Latreille, 1809) which are limits of these distributions were proposed and distributed in the entire Palaearctic region. The related to the contemporary and past develop- genera Camptopoeum Spinola, 1843 and Panur- ments which could have caused these ranges. gus Panzer, 1806 are also represented in the East- Palaearctic (in northern Thailand), but too few Catalogue of the old world species data are available for these genera to make them of Panurginus the subject of particular considerations. -

Plant Life MagillS Encyclopedia of Science
MAGILLS ENCYCLOPEDIA OF SCIENCE PLANT LIFE MAGILLS ENCYCLOPEDIA OF SCIENCE PLANT LIFE Volume 4 Sustainable Forestry–Zygomycetes Indexes Editor Bryan D. Ness, Ph.D. Pacific Union College, Department of Biology Project Editor Christina J. Moose Salem Press, Inc. Pasadena, California Hackensack, New Jersey Editor in Chief: Dawn P. Dawson Managing Editor: Christina J. Moose Photograph Editor: Philip Bader Manuscript Editor: Elizabeth Ferry Slocum Production Editor: Joyce I. Buchea Assistant Editor: Andrea E. Miller Page Design and Graphics: James Hutson Research Supervisor: Jeffry Jensen Layout: William Zimmerman Acquisitions Editor: Mark Rehn Illustrator: Kimberly L. Dawson Kurnizki Copyright © 2003, by Salem Press, Inc. All rights in this book are reserved. No part of this work may be used or reproduced in any manner what- soever or transmitted in any form or by any means, electronic or mechanical, including photocopy,recording, or any information storage and retrieval system, without written permission from the copyright owner except in the case of brief quotations embodied in critical articles and reviews. For information address the publisher, Salem Press, Inc., P.O. Box 50062, Pasadena, California 91115. Some of the updated and revised essays in this work originally appeared in Magill’s Survey of Science: Life Science (1991), Magill’s Survey of Science: Life Science, Supplement (1998), Natural Resources (1998), Encyclopedia of Genetics (1999), Encyclopedia of Environmental Issues (2000), World Geography (2001), and Earth Science (2001). ∞ The paper used in these volumes conforms to the American National Standard for Permanence of Paper for Printed Library Materials, Z39.48-1992 (R1997). Library of Congress Cataloging-in-Publication Data Magill’s encyclopedia of science : plant life / edited by Bryan D. -
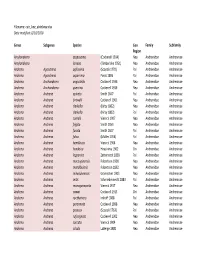
Anthophila List
Filename: cuic_bee_database. -

Effects of Habitat Fragmentation and Disturbance on Biodiversity and Ecosystem Functions
Effects of habitat fragmentation and disturbance on biodiversity and ecosystem functions Inauguraldissertation der Philosophisch-naturwissenschaftlichen Fakultät der Universität Bern vorgelegt von Christof Schüepp von Eschlikon TG Leiter der Arbeit: Prof. Dr. M. H. Entling Institut für Ökologie und Evolution | downloaded: 13.3.2017 Von der Philosophisch-naturwissenschaftlichen Fakultät angenommen. Bern, 14. Mai 2013 Der Dekan: Prof. Dr. S. Decurtins Originaldokument gespeichert auf dem Webserver der Universitätsbibliothek Bern Dieses Werk ist unter einem https://doi.org/10.7892/boris.54819 Creative Commons Namensnennung-Keine kommerzielle Nutzung-Keine Bearbeitung 2.5 Schweiz Lizenzvertrag lizenziert. Um die Lizenz anzusehen, gehen Sie bitte zu http://creativecommons.org/licenses/by-nc-nd/2.5/ch/ oder schicken Sie einen Brief an Creative Commons, 171 Second Street, Suite 300, San Francisco, California 94105, USA. source: Effects of habitat fragmentation and disturbance on biodiversity and ecosystem functions Creative Commons Licence Urheberrechtlicher Hinweis Dieses Dokument steht unter einer Lizenz der Creative Commons Namensnennung- Keine kommerzielle Nutzung-Keine Bearbeitung 2.5 Schweiz. http://creativecommons.org/licenses/by-nc-nd/2.5/ch/ Sie dürfen: dieses Werk vervielfältigen, verbreiten und öffentlich zugänglich machen Zu den folgenden Bedingungen: Namensnennung. Sie müssen den Namen des Autors/Rechteinhabers in der von ihm festgelegten Weise nennen (wodurch aber nicht der Eindruck entstehen darf, Sie oder die Nutzung des Werkes durch Sie würden entlohnt). Keine kommerzielle Nutzung. Dieses Werk darf nicht für kommerzielle Zwecke verwendet werden. Keine Bearbeitung. Dieses Werk darf nicht bearbeitet oder in anderer Weise verändert werden. Im Falle einer Verbreitung müssen Sie anderen die Lizenzbedingungen, unter welche dieses Werk fällt, mitteilen. Jede der vorgenannten Bedingungen kann aufgehoben werden, sofern Sie die Einwilligung des Rechteinhabers dazu erhalten. -

1. Padil Species Factsheet Scientific Name: Common Name Image
1. PaDIL Species Factsheet Scientific Name: Panurginus ineptus Cockerell, 1922 (Hymenoptera: Andrenidae: Panurginae: Panurgini) Common Name Tribe Representative - Panurgini Live link: http://www.padil.gov.au/pollinators/Pest/Main/139784 Image Library Australian Pollinators Live link: http://www.padil.gov.au/pollinators/ Partners for Australian Pollinators image library Western Australian Museum https://museum.wa.gov.au/ South Australian Museum https://www.samuseum.sa.gov.au/ Australian Museum https://australian.museum/ Museums Victoria https://museumsvictoria.com.au/ 2. Species Information 2.1. Details Specimen Contact: Museum Victoria - [email protected] Author: Ken Walker Citation: Ken Walker (2010) Tribe Representative - Panurgini(Panurginus ineptus)Updated on 8/11/2010 Available online: PaDIL - http://www.padil.gov.au Image Use: Free for use under the Creative Commons Attribution-NonCommercial 4.0 International (CC BY- NC 4.0) 2.2. URL Live link: http://www.padil.gov.au/pollinators/Pest/Main/139784 2.3. Facets Bio-Region: USA and Canada, Europe and Northern Asia Host Family: Not recorded Host Genera: Fresh Flowers Status: Exotic Species not in Australia Bio-Regions: Palaearctic, Nearctic Body Hair and Scopal location: Body hair relatively sparse, Tibia Episternal groove: Present but not extending below scrobal groove Wings: Submarginal cells - Three, Apex of marginal cell truncate or rounded Head - Structures: Two subantennal sutures below each antennal socket, Facial fovea present usually as a broad groove, Facial -

Hymenoptera, Apoidea)
>lhetian JMfuseum ox4tates PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK 24, N.Y. NUMBER 2 2 24 AUGUST I7, I 965 The Biology and Immature Stages of Melitturga clavicornis (Latreille) and of Sphecodes albilabris (Kirby) and the Recognition of the Oxaeidae at the Family Level (Hymenoptera, Apoidea) BYJEROME G. ROZEN, JR.' Michener (1944) divided the andrenid subfamily Panurginae into two tribes, the Panurgini and Melitturgini, with the latter containing the single Old World genus Melitturga. This genus was relegated to tribal status apparently on the grounds that the adults, unlike those of other panurgines, bear certain striking resemblances to the essentially Neo- tropical Oxaeinae of the same family. In 1951 Rozen showed that the male genitalia of Melitturga are unlike those of the Oxaeinae and are not only typical of those of the Panurginae in general but quite like those of the Camptopoeum-Panurgus-Panurginus-Epimethea complex within the sub- family. On the basis of this information, Michener (1954a) abandoned the idea that the genus Melitturga represents a distinct tribe of the Panur- ginae. Recently evidence in the form of the larva of Protoxaea gloriosa Fox (Rozen, 1965) suggested that the Oxaeinae were so unlike other Andreni- dae that they should be removed from the family unless some form inter- 1 Chairman and Associate Curator, Department of Entomology, the American Museum of Natural History. 2 AMERICAN MUSEUM NOVITATES NO. 2224 mediate between the two subfamilies is found. In spite of the structure of the male genitalia, Melitturga is the only known possible intermediary. -

Hymenoptera, Apidae, Biastini)
JHR 69: 23–37 (2019)Monotypic no more – a new species of the unusual genus Schwarzia... 23 doi: 10.3897/jhr.69.32966 RESEARCH ARTICLE http://jhr.pensoft.net Monotypic no more – a new species of the unusual genus Schwarzia (Hymenoptera, Apidae, Biastini) Silas Bossert1,2 1 Department of Entomology, Cornell University, Ithaca, New York 14853, USA 2 Department of Entomology, National Museum of Natural History, Smithsonian Institution, Washington, DC 20560, USA Corresponding author: Silas Bossert ([email protected]) Academic editor: Michael Ohl | Received 10 January 2019 | Accepted 21 March 2019 | Published 30 April 2019 http://zoobank.org/F30BBBB6-7C9C-44F0-A1C6-531C1521D3EC Citation: Bossert S (2019) Monotypic no more – a new species of the unusual genus Schwarzia (Hymenoptera, Apidae, Biastini). Journal of Hymenoptera Research 69: 23–37. https://doi.org/10.3897/jhr.69.32966 Abstract Schwarzia elizabethae Bossert, sp. n., a previously unknown species of the enigmatic cleptoparasitic genus Schwarzia Eardley, 2009 is described. Both sexes are illustrated and compared to the type species of the genus, Schwarzia emmae Eardley, 2009. The male habitus of S. emmae is illustrated and potential hosts of Schwarzia are discussed. Unusual morphological features of Schwarzia are examined in light of the pre- sumably close phylogenetic relationship to other Biastini. The new species represents the second species of Biastini outside the Holarctic region. Keywords Bees, cleptoparasitism, East Africa Introduction Biastini (Apidae, Nomadinae) is a small tribe of cleptoparasitic bees with just 13 de- scribed species in four genera (Ascher and Pickering 2019). Their distribution is pri- marily Holarctic. Biastes Panzer, 1806 is Palearctic and has five described species.