Understanding Antisocial Behavior Through Autonomic Arousal, Nutrition, and Non-Invasive Brain Stimulation
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Toward an Integrative Science of the Brain and Crime
BioSocieties https://doi.org/10.1057/s41292-019-00167-3 COMMENTARY Mind the gap: toward an integrative science of the brain and crime Eyal Aharoni1 · Nathaniel E. Anderson2 · J. C. Barnes3 · Corey H. Allen4 · Kent A. Kiehl2 © Springer Nature Limited 2019 In the recent article, Criminalizing the brain: Neurocriminology and the production of strategic ignorance, Fallin et al. (2018) argue that most neuroscientists working with antisocial populations are guilty of suppressing, obfuscating, and erasing legiti- mate social explanations for criminal behavior as part of a strategic attempt to tout their reductionist preconceptions of behavior and to advance their own professional, extra-scientifc agendas. While we applaud their call for greater inclusion of social/ contextual factors into neurocriminological research, we fnd that they overstate the case against neurocriminology and understate important eforts by this emerg- ing community to generate signifcant and cross-disciplinary contributions to the understanding of antisocial behavior. Learning to identify and avoid such mischar- acterizations is critical for the pursuit of much-needed interdisciplinary research and collaboration on the prediction, explanation, and remediation of antisocial behavior. By critically evaluating Fallin et al.’s claims, we hope to strike a conciliatory bal- ance, which is more likely to promote integration rather than antagonism between disciplines. Fallin and colleagues correctly describe biosocial criminology as an emerging discipline that integrates historically neglected biological factors and neuroscientifc methods in the study of antisocial behavior. Indeed, this paradigm has demonstrated the value of genetics (Brunner et al. 1993), physiology (Latvala et al. 2015), bio- chemistry (Coccaro et al. 1998), and neuroimaging (Anderson and Kiehl 2012) for * Eyal Aharoni [email protected] 1 Department of Psychology, Georgia State University, P.O. -

UNIT 9 CRIMINOLOGY THEORIES Criminological Theories
UNIT 9 CRIMINOLOGY THEORIES Criminological Theories Structure 9.1 Introduction 9.2 Objectives 9.3 The Study of Criminology 9.4 What is Criminology? 9.5 Brief History of Criminology 9.6 Classical School of Criminology 9.6.1 Pre Classical School 9.6.2 Classical School of Criminology 9.6.3 Neo Classical School 9.7 Positive School of Criminology 9.8 Ecological School of Criminology 9.9 Theories Related to Physical Appearance 9.9.1 Phsiognomy and Phrenology 9.9.2 Criminal Anthropology: Lombroso to Goring 9.9.3 Body Type Theories: Sheldon to Cortes 9.10 Biological Factors and Criminal Behaviour 9.10.1 Chromosomes and Crime 9.10.2 Family Studies 9.10.3 Twin and Adoption Studies 9.10.4 Neurotransmitters 9.10.5 Hormones 9.10.6 The Autonomic Nervous System 9.11 Psychoanalytical Theories of Crime 9.11.1 Psychanalytic Explanations of Criminal Behaviour 9.12 Sociological Theories of Criminal Behaviour 9.12.1 Durkhiem, Anomie and Modernisation 9.12.2 Merton’s Strain Theory 9.12.3 Sutherland’s Differential Association Theory 9.13 Critical Criminology 9.13.1 Marxim and Marxist Criminology 9.14 Control Theories 9.14.1 Drift and Neutralisation 9.14.2 Hirschi’s Social Control 9.14.3 Containment Theory 9.14.4 Labeling Theory 9.15 Conflict Theories 9.15.1 Sellin’s Culture Conflict Theory 9.15.2 Vold’s Group Conflict Theory 9.15.3 Quinney’s Theory of the Social Reality of Crime 9.15.4 Turk’s Theory of Criminalisation 9.15.5 Chambliss and Seidman’s Analysis of the Criminal Justice System 5 Theories and Perspectives 9.16 Summary in Criminal Justice 9.17 Terminal Questions 9.18 Answers and Hints 9.19 References and Suggested Readings 9.1 INTRODUCTION Criminology is the scientific approach towards studying criminal behaviour. -

The Growth of Criminological Theories
THE GROWTH OF CRIMINOLOGICAL THEORIES Jonathon M. Heidt B.A., University of Montana, 2000 THESIS SUBMITTED IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF MASTER OF ARTS In the School of Criminology OJonathon M. Heidt 2003 SIMON FRASER UNIVERSITY November 2003 All rights reserved. This work may not be reproduced in whole or in part, by photocopy or other means, without permission of the author. APPROVAL Name: Jonathon Heidt Degree: M.A. Title of Thesis: The Growth of Criminological Theories Examining Committee: Chair: ~ridnkurtch,P~JJ$ . D;. Robert ~ordoi,kh.~. Senior Supervisor Dr. Elizabeth Elliott, Ph.D. Member Sociology Department University at Albany - SUNY Date Approved: PARTIAL COPYRIGHT LICENCE I hereby grant to Simon Fraser University the right to lend my thesis, project or extended essay (the title of which is shown below) to users of the Simon Fraser University Library, and to make partial or single copies only for such users or in response to a request from the library of any other university, or other educational institution, on its own behalf or for one of its users. I further agree that permission for multiple copying of this work for scholarly purposes may be granted by me or the Dean of Graduate Studies. It is understood that copying or publication of this work for financial gain shall not be allowed without my written permission. Title of Thesis/Project/Extended Essay The Growth of Criminological Theories Author: Name ABSTRACT In the last 50 years, an extensive array of theories has appeared within the field of criminology, many generated by the discipline of sociology. -

Introduction to Criminology
PART 1 © Nevarpp/iStockphoto/Getty Images Introduction to Criminology CHAPTER 1 Crime and Criminology. 3 CHAPTER 2 The Incidence of Crime . 35 1 © Tithi Luadthong/Shutterstock CHAPTER 1 Crime and Criminology Crime and the fear of crime have permeated the fabric of American life. —Warren E. Burger, Chief Justice, U.S. Supreme Court1 Collective fear stimulates herd instinct, and tends to produce ferocity toward those who are not regarded as members of the herd. —Bertrand Russell2 OBJECTIVES • Define criminology, and understand how this field of study relates to other social science disciplines. Pg. 4 • Understand the meaning of scientific theory and its relationship to research and policy. Pg. 8 • Recognize how the media shape public perceptions of crime. Pg. 19 • Know the criteria for establishing causation, and identify the attributes of good research. Pg. 13 • Understand the politics of criminology and the importance of social context. Pg. 18 • Define criminal law, and understand the conflict and consensus perspectives on the law. Pg. 5 • Describe the various schools of criminological theory and the explanations that they provide. Pg. 9 of the public’s concern about the safety of their com- Introduction munities, crime is a perennial political issue that can- Crime is a social phenomenon that commands the didates for political office are compelled to address. attention and energy of the American public. When Dealing with crime commands a substantial por- crime statistics are announced or a particular crime tion of the country’s tax dollars. Criminal justice sys- goes viral, the public demands that “something be tem operations (police, courts, prisons) cost American done.” American citizens are concerned about their taxpayers over $270 billion annually. -

Sociological Theories of Deviance: Definitions & Considerations
Sociological Theories of Deviance: Definitions & Considerations NCSS Strands: Individuals, Groups, and Institutions Time, Continuity, and Change Grade level: 9-12 Class periods needed: 1.5- 50 minute periods Purpose, Background, and Context Sociologists seek to understand how and why deviance occurs within a society. They do this by developing theories that explain factors impacting deviance on a wide scale such as social frustrations, socialization, social learning, and the impact of labeling. Four main theories have developed in the last 50 years. Anomie: Deviance is caused by anomie, or the feeling that society’s goals or the means to achieve them are closed to the person Control: Deviance exists because of improper socialization, which results in a lack of self-control for the person Differential association: People learn deviance from associating with others who act in deviant ways Labeling: Deviant behavior depends on who is defining it, and the people in our society who define deviance are usually those in positions of power Students will participate in a “jigsaw” where they will become knowledgeable in one theory and then share their knowledge with the rest of the class. After all theories have been presented, the class will use the theories to explain an historic example of socially deviant behavior: Zoot Suit Riots. Objectives & Student Outcomes Students will: Be able to define the concepts of social norms and deviance 1 Brainstorm behaviors that fit along a continuum from informal to formal deviance Learn four sociological theories of deviance by reading, listening, constructing hypotheticals, and questioning classmates Apply theories of deviance to Zoot Suit Riots that occurred in the 1943 Examine the role of social norms for individuals, groups, and institutions and how they are reinforced to maintain a order within a society; examine disorder/deviance within a society (NCSS Standards, p. -

Neuroscience, Law, and Ethics T
International Journal of Law and Psychiatry 65 (2019) 101459 Contents lists available at ScienceDirect International Journal of Law and Psychiatry journal homepage: www.elsevier.com/locate/ijlawpsy Introduction Neuroscience, Law, and Ethics T In his influential Hardwired Behavior: What Neuroscience Reveals concerning the relevance of neurosciences to the law, especially crim- about Morality (2005), Laurence Tancredi combined neuroscience, inal law.” (Meynen, 2016, p. 3, Meynen, 2014; Morse & Roskies, 2013; psychology, philosophy, legal theory and clinical cases to insightfully Glenn & Raine, 2014; Chandler, 2018). One example is the use of examine the role of brain structure and function in moral behavior. neuroimaging to support behavioral evidence in judging whether a Analyzing how neurobiology shapes reasoning and decision-making, he person performing a criminal act met cognitive and volitional condi- discussed questions such as whether or to what extent we have free will tions necessary for criminal responsibility. Functional imaging re- and can be morally and criminally responsible for our actions. Is our cording abnormal activity in prefrontal-limbic pathways may indicate behavior hardwired and thus determined by the brain? Is free will an impaired cognitive and emotional processing and impaired capacity to illusion? Can mental content be explained entirely in terms of neural deliberate and consider the consequences of what one does or fails to content? If the brain influences but does not determine behavior, then do. This and other forms of imaging may eventually offer a more fine- how much of what we think and do is up to us? In his new book, grained account of impaired inhibitory mechanisms in the brain. -

The New BRAIN
SPECIAL EDITION The New BRAIN MACLEAN’S EBOOK Contents Introduction Join us for a giant brainstorming session on what the world’s neuroscience superstars are keeping top of mind The glia club Once dismissed as ‘glue,’ glial cells, neuron’s little brother, have become the lodestone of brain research. But is it a good idea for scientists to herd in one direction? Charlie Gillis How to build a brain A philosopher and engineer has created the most complex simulated brain in the world. On $30,000 a year. Nick Taylor-Vaisey Mad beauty A conceptual photographer dusts off the jars of a brain collection from a Texas mental hospital David Graham They grow up so fast The latest research on a baby’s remarkable brain development, from recognizing right and wrong to the gift of memory Rosemary Counter Gone baby gone Why don’t we remember anything from earliest childhood? It’s called infantile amnesia. Emma Teitel MACLEAN’S EBOOK THE NEW BRAIN Memory and gender Emma Teitel The young and the restless No one knows why autistic kids are often night owls, but their parents can take heart: science is looking at some biological causes based in the brain Katherine DeClerq Crying out for attention How one psychologist is offering hope to parents worried about the stigma, safety and side effects of ADHD medication Hannah Hoag Mind the age gap Previously dismissed as lesser or defective, new research is revealing that the teenage brain is just as powerful as any adult’s Rosemary Counter No brawn, no brains Genetics may decide your upper and lower limits for cognitive -

Social Structure, Culture, and Crime: Assessing Kornhauser’S Challenge to Criminology1
6 Social Structure, Culture, and Crime: Assessing Kornhauser’s Challenge to Criminology1 Ross L. Matsueda Ruth Kornhauser’s (1978) Social Sources of Delinquency has had a lasting infl uence on criminological theory and research. This infl uence consists of three contributions. First, Kornhauser (1978) developed a typology of criminological theories—using labels social disorganization, control, strain, and cultural devi- ance theories—that persists today. She distinguished perspectives by assump- tions about social order, motivation, determinism, and level of explanation, as well as by implications for causal models of delinquency. Second, Kornhauser addressed fundamental sociological concepts, including social structure, culture, and social situations, and provided a critical evaluation of their use within differ- ent theoretical perspectives. Third, she helped foster a resurgence of interest in social control and social disorganization theories of crime (e.g., Gottfredson and Hirschi 1990; Hirschi 1969; Sampson and Laub 1993; Bursik and Webb 1982; Kubrin and Weitzer 2003; Sampson and Groves 1989), both indirectly via the infl uence of an early version of the manuscript that would become Social Sources (see Chapter 3 in this volume) on Hirschi’s (1969) Causes of Delinquency , and directly via the infl uence of her book on the revitalization of social disorgani- zation theories of crime. Kornhauser’s infl uence on the writings of Hirschi, Bursik, and Sampson was particularly signifi cant and positive, as control and disorganization theo- ries came to dominate the discipline, stimulating a large body of research that increased our understanding of conventional institutional control of crime and delinquency through community organization, family life, and school effects. -

Tanksley, J.C
Identifying psychological pathways to polyvictimization: evidence from a longitudinal cohort study of twins from the UK Peter T. Tanksley, J.C. Barnes, Brian B. Boutwell, Louise Arseneault, Avshalom Caspi, Andrea Danese, Helen L. Fisher, et al. Journal of Experimental Criminology ISSN 1573-3750 J Exp Criminol DOI 10.1007/s11292-020-09422-1 1 23 Your article is protected by copyright and all rights are held exclusively by Springer Nature B.V.. This e-offprint is for personal use only and shall not be self-archived in electronic repositories. If you wish to self-archive your article, please use the accepted manuscript version for posting on your own website. You may further deposit the accepted manuscript version in any repository, provided it is only made publicly available 12 months after official publication or later and provided acknowledgement is given to the original source of publication and a link is inserted to the published article on Springer's website. The link must be accompanied by the following text: "The final publication is available at link.springer.com”. 1 23 Author's personal copy Journal of Experimental Criminology https://doi.org/10.1007/s11292-020-09422-1 Identifying psychological pathways to polyvictimization: evidence from a longitudinal cohort study of twins from the UK Peter T. Tanksley1 & J.C. Barnes1 & Brian B. Boutwell2,3 & Louise Arseneault4 & Avshalom Caspi4,5,6,7 & Andrea Danese4,8 & Helen L. Fisher4 & Terrie E. Moffitt4,5,6,7 # Springer Nature B.V. 2020 Abstract Objectives Examine the extent to which cognitive/psychological characteristics predict later polyvictimization. We employ a twin-based design that allows us to test the social neurocriminology hypothesis that environmental factors influence brain-based charac- teristics and influence behaviors like victimization. -
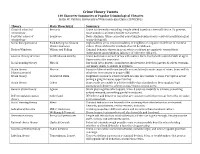
Crime Theory Tweets 140 Character Summaries of Popular Criminological Theories Justin W
Crime Theory Tweets 140 Character Summaries of Popular Criminological Theories Justin W. Patchin, University of Wisconsin-Eau Claire (CRMJ 301) Theory Main Theorist(s) Summary Classical school of Beccaria Crime is inherently rewarding. People offend based on a free will choice. To prevent, criminology must punish so potential benefit not worth it. Positivist school of Lombroso Born criminals. Crime caused by something beyond person’s control (usually biological criminology or psychological). Social disorganization Park & Burgess; Shaw & High mobility areas result in inability of neighbors to organize in defense of common McKay; Sampson values. Physical disorder symbols of social breakdown. Broken Windows Wilson and Kelling Criminal behavior thrives in areas where residents are apathetic toward their environment and neighbors (absence of collective efficacy). General theory of crime Gottfredson & Hirschi Crime & deviance a result of low self-control. One’s level of self-control stabile at age 8. Opportunity also important. Social bonding theory Hirschi Our bond (attachment, commitment, involvement, belief) to parents & others restrains our innate desire to engage in deviance. Strain theory Merton Pursuit of American Dream (wealth accumulation) is main cause of crime. Some will do (classic/anomie) whatever is necessary to acquire $$$. Strain theory Cloward & Ohlin Illegitimate means to achieve wealth are also inaccessible to some. Perception is that joining a gang increases opportunities. Strain theory Cohen Some youth are unable to achieve middle-class standards so they supplant legit pursuits with desire to achieve status/respect among peers. General strain theory Agnew Strain plus negative affect equals crime. 3 sources: inability to achieve; something valued removed; something painful introduced. -

Neuroimaging Abnormalities in Neurological Patients with Criminal Behavior
Current Neurology and Neuroscience Reports (2018) 18:47 https://doi.org/10.1007/s11910-018-0853-3 BEHAVIOR (H S KIRSHNER, SECTION EDITOR) Neuroimaging Abnormalities in Neurological Patients with Criminal Behavior R. Ryan Darby1 # Springer Science+Business Media, LLC, part of Springer Nature 2018 Abstract Purpose of Review Criminal behavior occurs in previously law-abiding neurological patients, including patients with traumatic brain injury, focal brain lesions, and dementia. Neuroimaging abnormalities in these patients allow one to explore the potential neuroanatomical correlates of criminal behavior. However, this process has been challenging because (1) It is difficult to determine the temporal relationship between criminal behavior and neurological disease onset; (2) Abnormalities in several different brain regions have been associated with criminal behavior; and (3) It is difficult to quantify neuroimaging abnormalities in individual subjects. Recent Findings Recent studies have begun to address these concerns, showing that neuroimaging abnormalities in patients with criminal behavior localize to a common brain network, rather than a single specific brain region. New methods have been developed to identify atrophy patterns in individual patients, but have not yet been used in neurological patients with criminal behavior. Summary Future advances will be important for making sure that neuroimaging data is used in a responsible manner in legal cases involving criminal behavior. Keywords Criminal . Moral . Brain lesions . Neurology . Frontal lobe . Brain networks Introduction he was killed by police [2]. At autopsy, he was found to have a malignant tumor in his right temporal lobe, prompting ques- Neurological diseases can cause profound changes to behav- tions about the degree to which his brain injury contributed to ior. -

Readings for Graduate Criminology Comprehensive Exam
READINGS FOR GRADUATE CRIMINOLOGY COMPREHENSIVE EXAM The following subdivisions are simply meant to be a heuristic device. They are not necessarily mutually exclusive or exhaustive. Note: Students are responsible for the last 5 years of articles in scholarly journals related to the area including Criminology, Criminology and Public Policy, and related papers published in the American Sociological Review, the American Journal of Sociology, and Social Forces and all other top journals. Social Disorganization and the Chicago School Bursik, R.J. 1988. "Social Disorganization theories of crime and delinquency: Problems and Prospects." Criminology 26:519-552. Morenoff, J., R. Sampson and S. Raudenbush. 2001. “Neighborhood inequality, collective efficacy, and the spatial dynamics of urban violence.” Criminology 39 (3): 517-559. Park, Robert E. 1915. “The City: Suggestions for the Investigation of Human Behavior in the City Environment. American Journal of Sociology 20: 577-612. Rose, D. and T. Clear. 1998. "Incarceration, social capital and crime: implications for social disorganization theory." Criminology 36:441-479. Sampson, R. 2012. The Great American City: Chicago and the Enduring Neighborhood Effect. University of Chicago Press. Sampson, R., S.W. Raudenbush and F. Earls. 1997."Neighborhoods and violent crime: A multilevel study of collective efficacy." Science 277: 918-924. Sampson, R., and Groves, W.B. (1989). “Community Structure and Crime: Testing Social- Disorganization Theory.” American Journal of Sociology 94 (4): 774-802. Shaw, Clifford R., and Henry D. McKay. 1942. Juvenile Delinquency in Urban Areas. Chicago: University of Chicago Press. Differential Association and Social Learning Theories Akers, R. L. 1998. Social Learning and Social Structure: A General Theory of Crime and Deviance.