Dormancy and Germination
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Plant Physiology
PLANT PHYSIOLOGY Vince Ördög Created by XMLmind XSL-FO Converter. PLANT PHYSIOLOGY Vince Ördög Publication date 2011 Created by XMLmind XSL-FO Converter. Table of Contents Cover .................................................................................................................................................. v 1. Preface ............................................................................................................................................ 1 2. Water and nutrients in plant ............................................................................................................ 2 1. Water balance of plant .......................................................................................................... 2 1.1. Water potential ......................................................................................................... 3 1.2. Absorption by roots .................................................................................................. 6 1.3. Transport through the xylem .................................................................................... 8 1.4. Transpiration ............................................................................................................. 9 1.5. Plant water status .................................................................................................... 11 1.6. Influence of extreme water supply .......................................................................... 12 2. Nutrient supply of plant ..................................................................................................... -

What Are Soybeans?
candy, cakes, cheeses, peanut butter, animal feeds, candles, paint, body lotions, biodiesel, furniture soybeans USES: What are soybeans? Soybeans are small round seeds, each with a tiny hilum (small brown spot). They are made up of three basic parts. Each soybean has a seed coat (outside cover that protects the seed), VOCABULARY cotyledon (the first leaf or pair of leaves within the embryo that stores food), and the embryo (part of a seed that develops into Cultivar: a variety of plant that has been created or a new plant, including the stem, leaves and roots). Soybeans, selected intentionally and maintained through cultivation. like most legumes, perform nitrogen fixation. Modern soybean Embryo: part of a seed that develops into a new plant, cultivars generally reach a height of around 1 m (3.3 ft), and including the stem, leaves and roots. take 80–120 days from sowing to harvesting. Exports: products or items that the U.S. sells and sends to other countries. Exports include raw products like whole soybeans or processed products like soybean oil or Leaflets soybean meal. Fertilizer: any substance used to fertilize the soil, especially a commercial or chemical manure. Hilum: the scar on a seed marking the point of attachment to its seed vessel (the brown spot). Leaflets: sub-part of leaf blade. All but the first node of soybean plants produce leaves with three leaflets. Legume: plants that perform nitrogen fixation and whose fruit is a seed pod. Beans, peas, clover and alfalfa are all legumes. Nitrogen Fixation: the conversion of atmospheric nitrogen Leaf into a nitrogen compound by certain bacteria, such as Stem rhizobium in the root nodules of legumes. -

African Mahogany Anegre Birdseye Maple Black Walnut
African Mahogany African Mahogany (Khaya) is a genus of seven species of trees in the mahogany family Meliaceae, native to tropical Africa and Madagascar. All species become big trees 30-35 m tall, rarely 45 m, with a trunk over 1 m trunk diameter, often buttressed at the base. The leaves are pinnate, with 4-6 pairs of leaflets, the terminal leaflet absent; each leaflet is 10-15 cm long abruptly rounded toward the apex but often with an acuminate tip. The leaves can be either deciduous or evergreen depending on the species. The flowers are produced in loose inflorescences, each flower small, with four or five yellowish petals and ten stamens. The timber of Khaya is called African mahogany, the only timber widely accepted as mahogany besides that of the true mahogany, of the genus Swietenia. Khaya senegalensis, also known as the African dry zone mahogany is also used for its non timber parts. In West Africa, Fulani herdsmen prune the tree during the dry season to feed cattle. Anegre Anegre is milled from the Tawa tree (Beilschmiedia tawa) is a New Zealand broadleaf tree common in the central parts of the country. Tawa is often the dominant canopy species in lowland forests in the North Island and north east of the South Island, Individual specimens may grow up to 30 meters or more in height with trunks up to 1.2 meters in diameter, and they have smooth dark bark. The word "tawa" is the Maori name for the tree. One of the few hardwood trees in the country with good timber, the wood of this tree can be used for attractive and resilient floor boarding. -

International Union for the Protection of New Varieties of Plants
E TG/81/7(proj.1) ORIGINAL: English DATE: 2018-04-05 INTERNATIONAL UNION FOR THE PROTECTION OF NEW VARIETIES OF PLANTS Geneva DRAFT * COMMON SUNFLOWER UPOV Code(s): HLNTS_ANN Helianthus annuus L. GUIDELINES FOR THE CONDUCT OF TESTS FOR DISTINCTNESS, UNIFORMITY AND STABILITY prepared by experts from Hungary to be considered by the Technical Working Party for Agricultural Crops at its forty-seventh session, to be held in Naivasha, Kenya, from 2018-05-21 to 2018-05-25 Disclaimer: this document does not represent UPOV policies or guidance Alternative names:* Botanical name English French German Spanish Helianthus annuus L. Common Sunflower Soleil, Tournesol Sonnenblume Girasol The purpose of these guidelines (“Test Guidelines”) is to elaborate the principles contained in the General Introduction (document TG/1/3), and its associated TGP documents, into detailed practical guidance for the harmonized examination of distinctness, uniformity and stability (DUS) and, in particular, to identify appropriate characteristics for the examination of DUS and production of harmonized variety descriptions. ASSOCIATED DOCUMENTS These Test Guidelines should be read in conjunction with the General Introduction and its associated TGP documents. * These names were correct at the time of the introduction of these Test Guidelines but may be revised or updated. [Readers are advised to consult the UPOV Code, which can be found on the UPOV Website (www.upov.int), for the latest information.] TG/81/7(proj.1) Common Sunflower, 2018-04-05 2 TABLE OF CONTENTS PAGE 1. -

Notes on the Genus Ormosia (Fabaceae-Sophoreae) in Thailand
THAI FOREST BULL., BOT. 45(2): 118–124. 2017. DOI https://doi.org/10.20531/tfb.2017.45.2.07 Notes on the genus Ormosia (Fabaceae-Sophoreae) in Thailand SAWAI MATTAPHA1,*, SOMRAN SUDDEE2 & SUKID RUEANGRUEA2 ABSTRACT Ormosia mekongensis Mattapha, Suddee & Rueangr. is described as a new species and illustrated. Its conservation status is assessed and its distribution is mapped. Three other species, Ormosia grandistipulata Whitmore, O. penangensis Ridl. and O. venosa Baker, are updated for the generic account for the Flora of Thailand: the first could now be fully described, because flowers were found, the latter two are new records for peninsular Thailand. KEYWORDS: Lectotypifications, Mekong, new species, new record, PeninsularThailand. Published online: 1 December 2017 INTRODUCTION Niyomdham, Thai Forest Bull., Bot. 13: 5, f. 2. 1980. Type: Malaysia, Trengganu, 1955, Sinclair & Kiah Ormosia Jacks., a genus in the tribe Sophoreae bin Salleh SFN 40851 (holotype SING; isotypes K!, of the Leguminosae, comprises approximately 90 L!-digital images). species distributed in Asia, the Americas and Australia (Queensland) (Lewis et al., 2005). The Tree 10–15 m tall; young shoots, inflorescences genus was revised for Thailand by Niyomdham and calyces pubescent. Leaves: rachis 20–25 cm (1980), who accepted eight indigenous species, and long, apex acute, puberulous; petioles 6–10 cm long; here we add three species, which bring the total stipules large, ovate, 2–5 by 1–3 cm, puberulous on number of Ormosia species for the Flora of Thailand both sides, persistent; leaflets 9–13, coriaceous, account to 11. The three species that are new to oblong-obovate, 4–18 by 2.5–8 cm, upper surface Thailand are O. -

Effects of Root Pruning on Germinated Pecan Seedlings
PROPAGATION AND TISSUE CULTURE HORTSCIENCE 50(10):1549–1552. 2015. if pruning the roots shortly after germination stimulates lateral root formation. Similarly, little work has been attempted to consider Effects of Root Pruning on Germinated the effects of taproot pruning in young pecan seedlings back to different lengths on root Pecan Seedlings regeneration. The objective of this study 1 was to determine if root pruning at different Rui Zhang, Fang-Ren Peng , Pan Yan, Fan Cao, periods after seed germination affects pecan and Zhuang-Zhuang Liu seedling root and shoot growth and to evaluate College of Forestry, Nanjing Forestry University, 159 Longpan Road, different degrees of taproot pruning on root Nanjing 210037, China initiation. Dong-Liang Le Materials and Methods College of Forestry, Nanjing Forestry University, 159 Longpan Road, Nanjing 210037, China; and Nanjing Green Universe Pecan Science and The test was established at Lvzhou Pecan Research Station, Nanjing, China (lat. Technology Co. Ltd., 38 Muxuyuan Street, Nanjing 210007, China 32.05°N, long. 118.77°E). Open-pollinated Peng-Peng Tan seeds of the Chinese selection ‘Shaoxing’ were used as seed stock to produce seedlings. College of Forestry, Nanjing Forestry University, 159 Longpan Road, ‘Shaoxing’ had a small nut of very good Nanjing 210037, China quality with 50% percentage fill. The seeds averaged 30.4 mm in length, 20.9 mm in Additional index words. Carya illinoinensis, taproot, lateral root, shoot growth width, and 5.4 g in weight. Seeds were Abstract. Root systems of pecan trees are usually dominated by a single taproot with few collected on 7 Nov. -

African Mahogany Wood Defects Detected by Ultrasound Waves
General Technical Report FPL-GTR-239 • Proceedings: 19th International Nondestructive Testing and Evaluation of Wood Symposium African mahogany wood defects detected by ultrasound waves Tamara Suely Filgueira Amorim França Department of Sustainable Bioproducts, Mississippi State University, Starkville, Mississippi, United States, [email protected] Frederico Jose Nistal França Department of Sustainable Bioproducts, Mississippi State University, Starkville, Mississippi, United States, [email protected] Robert John Ross U.S. Forest Service, Forest Product Laboratory, Madison, Wisconsin, USA, [email protected] Xiping Wang U.S. Forest Service, Forest Product Laboratory, Madison, Wisconsin, USA, [email protected] Marina Donaria Chaves Arantes Departamento de Ciências Florestais e da Madeira, Universidade Federal do Espírito Santo, Jerônimo Monteira, Espírito Santo, Brasil, [email protected] Roy Daniel Seale Department of Sustainable Bioproducts, Mississippi State University, Starkville, Mississippi, United States, [email protected] Abstract This study aims to investigate the potential of ultrasound wave to detect defects in 19 years old of two species of African mahogany planted in Brazil. Were used five 76 x 5 x 5 cm samples from each species with different types of defects, and were conditioned to 12% moisture content. The samples were scanned with ultrasound wave in longitudinal direction and every 2,54 cm in radial and tangential directions along the samples. It was possible to identify end split and pin knots in Khaya ivorensis and reaction wood in Khaya senegalensis wood. Beetle galleries did not affect wave velocities in Khaya senegalensis wood. Grain angle had a large effect in ultrasound velocities in radial and tangential directions. Khaya senegalensis exhibit lower longitudinal velocities related to larger amount of interlocked grain in this species. -

Lumber / Veneer Match
Lumber Veneer Type of Cut Alder (Select) Alder Plain Sliced Face Alder (Knotty) Knotty Alder (Dark Putty in open knots) Plain Sliced Face Ash (Eastern White) Ash (Eastern White) Rotary Multi Piece Face Ash (Western) Ash (Eastern White) Rotary Multi Piece Face Beech (European Steamed) Beech (European Steamed) Plain Sliced Face Birch (Eastern Red) N/A N/A Birch (Eastern White) Birch (Eastern White) Rotary Multi Piece Face Birch (Western) N/A Cherry (Select) Cherry (Select) Plain Sliced Face Cherry (Rustic) Fir (CVG) Fir (CVG) Plain Sliced Face Hickory (Brown Heart) Hickory (Brown Heart) Plain Sliced Face Hickory (Calico) Hickory (Calico) Plain Sliced Face Hickory (Rustic Brown Heart) Hickory (Rustic Brown Heart) Plain Sliced Face Hickory (Rustic Calico) Hickory (Rustic Calico) Plain Sliced Face Lyptus Lyptus Plain Sliced Face Mahogany (African Khaya) Mahogany (African Khaya) Plain Sliced Face Mahogany (Edinam) Mahogany (African Khaya) Plain Sliced Face Maple (Eastern Hard White) Maple (Eastern Hard White) Rotary Whole Piece Face Maple (Eastern Soft Rustic) Maple (Eastern Hard Rustic) Plain Sliced Face Maple (Eastern Soft White) Maple (Eastern Hard White) Rotary Whole Piece Face Maple (Western) Alder Plain Sliced Face Paint Grade (Lumber Panel) DCD's Choice Rotary Whole Piece Face Paint Grade (MDF Panel) MDF N/A Pine (Eastern Clear) Pine (Eastern Clear) Plain Sliced Face Pine (Eastern Knotty) Pine (Eastern Knotty) Plain Sliced Face Poplar N/A N/A Red Oak (Natural) Red Oak (Natural) Plain Sliced Face Red Oak (Qtr Sawn) Red Oak (Qtr Sawn) Quarter Sawn Face Red Oak (Rift) Red Oak (Rift) Rift Sawn Face Red Oak (Select) Red Oak (Select) Plain Sliced Face Sapele (Flat Sawn) Sapele (Flat Sawn) Plain Sliced Face Sapele (Qtr Sawn) Sapele (Qtr Sawn) Quarter Sawn Face Walnut Walnut Plain Sliced Face White Oak (Select) White Oak (Select) Plain Sliced Face White Oak (Qtr Sawn) White Oak (Qtr Sawn) Quarter Sawn Face White Oak (Rift) White Oak (Rift) Rift Sawn Face. -

Chec List What Survived from the PLANAFLORO Project
Check List 10(1): 33–45, 2014 © 2014 Check List and Authors Chec List ISSN 1809-127X (available at www.checklist.org.br) Journal of species lists and distribution What survived from the PLANAFLORO Project: PECIES S Angiosperms of Rondônia State, Brazil OF 1* 2 ISTS L Samuel1 UniCarleialversity of Konstanz, and Narcísio Department C.of Biology, Bigio M842, PLZ 78457, Konstanz, Germany. [email protected] 2 Universidade Federal de Rondônia, Campus José Ribeiro Filho, BR 364, Km 9.5, CEP 76801-059. Porto Velho, RO, Brasil. * Corresponding author. E-mail: Abstract: The Rondônia Natural Resources Management Project (PLANAFLORO) was a strategic program developed in partnership between the Brazilian Government and The World Bank in 1992, with the purpose of stimulating the sustainable development and protection of the Amazon in the state of Rondônia. More than a decade after the PLANAFORO program concluded, the aim of the present work is to recover and share the information from the long-abandoned plant collections made during the project’s ecological-economic zoning phase. Most of the material analyzed was sterile, but the fertile voucher specimens recovered are listed here. The material examined represents 378 species in 234 genera and 76 families of angiosperms. Some 8 genera, 68 species, 3 subspecies and 1 variety are new records for Rondônia State. It is our intention that this information will stimulate future studies and contribute to a better understanding and more effective conservation of the plant diversity in the southwestern Amazon of Brazil. Introduction The PLANAFLORO Project funded botanical expeditions In early 1990, Brazilian Amazon was facing remarkably in different areas of the state to inventory arboreal plants high rates of forest conversion (Laurance et al. -
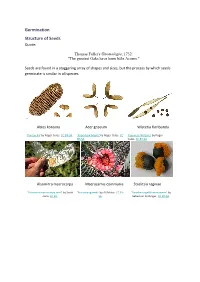
Germination Structure of Seeds Quote
Germination Structure of Seeds Quote: Thomas Fuller's Gnomologia, 1732: "The greatest Oaks have been little Acorns." Seeds are found in a staggering array of shapes and sizes, but the process by which seeds germinate is similar in all species. Abies koreana Acer griseum Wisteria floribunda 'Korean Fir' by Roger Culos. CC BY-SA. 'Paperbark Maple' by Roger Culos. CC 'Japanese Wisteria' by Roger BY-SA. Culos. CC BY-SA. Alsomitra macrocarpa Macrozamia communis Strelitzia reginae 'Alsomitra macrocarpa seed' by Scott 'BurrawangSeeds' by AYArktos. CC BY- 'Paradiesvogelblumensamen' by Zona. CC BY. SA. Sebastian Stabinger. CC BY-SA. Taraxicum officinale Stephanotis floribunda Phleum pratense 'Achane of Taraxacum sect. 'Stephanotis seed' by L. Marie"/Lenore 'Timoteegras vruchten Phleum Ruderalia' by Didier Edman, Sunnyvale, CA. CC BY. pratense' by Rasbak. CC BY-SA. Descouens. CC BY-SA. Dicotyledon seeds testa epicotyl plumule hypocotyl cotyledon radicle 'Aesculus hippocastanum seed section' by Boronian. CC BY. plumule epicotyl hypocotyl testa hilum radicle cotyledon micropyle endosperm Monocotyledon seeds endosperm epicotyl testa hypocotyl cotyledon radicle Parts of a seed Testa The seed coat. A protective layer which is tough and hard and it protects the seed from attack by insects, fungi and bacteria. Cotyledon Dicotyledons have 2 cotyledons Monocotyledons have 1 cotyledon A cotyledon is an embryonic leaf. It is the first leaf to appear when a seedling grows. They often contain reserves of food which the developing seedling can use to grow. Epicotyl The section of stem between the cotyledon(s) and the plumule. In a seedling it is the section of stem between the cotyledons and the first true leaves. -

Fruits and Seeds of Genera in the Subfamily Faboideae (Fabaceae)
Fruits and Seeds of United States Department of Genera in the Subfamily Agriculture Agricultural Faboideae (Fabaceae) Research Service Technical Bulletin Number 1890 Volume I December 2003 United States Department of Agriculture Fruits and Seeds of Agricultural Research Genera in the Subfamily Service Technical Bulletin Faboideae (Fabaceae) Number 1890 Volume I Joseph H. Kirkbride, Jr., Charles R. Gunn, and Anna L. Weitzman Fruits of A, Centrolobium paraense E.L.R. Tulasne. B, Laburnum anagyroides F.K. Medikus. C, Adesmia boronoides J.D. Hooker. D, Hippocrepis comosa, C. Linnaeus. E, Campylotropis macrocarpa (A.A. von Bunge) A. Rehder. F, Mucuna urens (C. Linnaeus) F.K. Medikus. G, Phaseolus polystachios (C. Linnaeus) N.L. Britton, E.E. Stern, & F. Poggenburg. H, Medicago orbicularis (C. Linnaeus) B. Bartalini. I, Riedeliella graciliflora H.A.T. Harms. J, Medicago arabica (C. Linnaeus) W. Hudson. Kirkbride is a research botanist, U.S. Department of Agriculture, Agricultural Research Service, Systematic Botany and Mycology Laboratory, BARC West Room 304, Building 011A, Beltsville, MD, 20705-2350 (email = [email protected]). Gunn is a botanist (retired) from Brevard, NC (email = [email protected]). Weitzman is a botanist with the Smithsonian Institution, Department of Botany, Washington, DC. Abstract Kirkbride, Joseph H., Jr., Charles R. Gunn, and Anna L radicle junction, Crotalarieae, cuticle, Cytiseae, Weitzman. 2003. Fruits and seeds of genera in the subfamily Dalbergieae, Daleeae, dehiscence, DELTA, Desmodieae, Faboideae (Fabaceae). U. S. Department of Agriculture, Dipteryxeae, distribution, embryo, embryonic axis, en- Technical Bulletin No. 1890, 1,212 pp. docarp, endosperm, epicarp, epicotyl, Euchresteae, Fabeae, fracture line, follicle, funiculus, Galegeae, Genisteae, Technical identification of fruits and seeds of the economi- gynophore, halo, Hedysareae, hilar groove, hilar groove cally important legume plant family (Fabaceae or lips, hilum, Hypocalypteae, hypocotyl, indehiscent, Leguminosae) is often required of U.S. -

Abstract Germinación De Semillas De Ormosia Macrocalyx, Un Árbol
GEORGINA VARGAS-SIMÓN1,2, PABLO MARTÍNEZ-ZURIMENDI1,3*, MARIVEL DOMÍNGUEZ-DOMÍNGUEZ4 AND REINALDO PIRE5 Botanical Sciences 95 (2): 329-341, 2017 Abstract Background: Ormosia macrocalyx is a tropical forest tree classified as endangered. Its seeds experience problems of dispersion and apparent physical dormancy due to their hard seed coating. DOI: 10.17129/botsci.823 Hypotheses: 1) The stages of dehiscence of the fruits of Ormosia macrocalyx influence the germinative behavior of its seeds. 2) Pregerminative treatments will improve the germination process of the seeds stored under refrigeration. Copyright: © 2017 Vargas-Simón et Study species: Ormosia macrocalyx al. This is an open access article dis- Study site and period: Villahermosa, Tabasco, Mexico from October 2012 to October 2014 tributed under the terms of the Crea- tive Commons Attribution License, Methods: Two completely randomized experiments were conducted in order to analyze the germination process by 1) which permits unrestricted use, dis- evaluating germination in seeds from fruit at three stages of dehiscence (closed, semi-open, and open fruit) and 2) ap- tribution, and reproduction in any plying four treatments, including three pregerminative treatments (water soaking for 24 h, mechanical scarification and medium, provided the original author scarification+1% gibberellic acid) plus an untreated control, to seeds stored under refrigeration for 17 months. and source are credited. Results: Differences were found in germination rate (GR), corrected germination rate (CGR) and time to attain 50 % ger- mination (T50) among treatments in the first experiment, with the seeds from open fruits presenting the lowest response -1 (3.31 % day for GR and CGR, and 15.8 days for T50), although all treatments showed similar times for the initiation Author Contributions of germination (GI) and final germination percentage (GP).