Humidity Levels in Pied Flycatcher Nests Measured Using Capsule Hygrometers
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

2 Powys Local Development Plan Written Statement
Powys LDP 2011-2026: Deposit Draft with Focussed Changes and Further Focussed Changes plus Matters Arising Changes September 2017 2 Powys Local Development Plan 2011 – 2026 1/4/2011 to 31/3/2026 Written Statement Adopted April 2018 (Proposals & Inset Maps published separately) Adopted Powys Local Development Plan 2011-2026 This page left intentionally blank Cyngor Sir Powys County Council Adopted Powys Local Development Plan 2011-2026 Foreword I am pleased to introduce the Powys County Council Local Development Plan as adopted by the Council on 17th April 2017. I am sincerely grateful to the efforts of everyone who has helped contribute to the making of this Plan which is so important for the future of Powys. Importantly, the Plan sets out a clear and strong strategy for meeting the future needs of the county’s communities over the next decade. By focussing development on our market towns and largest villages, it provides the direction and certainty to support investment and enable economic opportunities to be seized, to grow and support viable service centres and for housing development to accommodate our growing and changing household needs. At the same time the Plan provides the protection for our outstanding and important natural, built and cultural environments that make Powys such an attractive and special place in which to live, work, visit and enjoy. Our efforts along with all our partners must now shift to delivering the Plan for the benefit of our communities. Councillor Martin Weale Portfolio Holder for Economy and Planning -
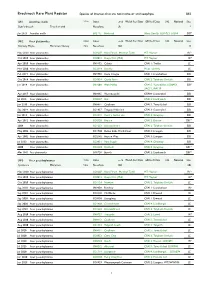
Brecknock Rare Plant Register Species of Interest That Are Not Native Or Archaeophyte S8/1
Brecknock Rare Plant Register Species of interest that are not native or archaeophyte S8/1 S8/1 Acanthus mollis 270m Status Local Welsh Red Data GB Red Data S42 National Sites Bear's-breech Troed yr arth Neophyte LR 1 Jun 2013 Acanthus mollis SO2112 Blackrock Mons: Llanelly: SSSI0733, SAC08 DB⁴ S8/2 Acer platanoides 260m Status Local Welsh Red Data GB Red Data S42 National Sites Norway Maple Masarnen Norwy 70m Neophyte NLS 18 Nov 2020 Acer platanoides SO0207 Nant Ffrwd, Merthyr Tydfil MT: Vaynor IR¹⁰ Oct 2020 Acer platanoides SO0012 Llwyn Onn (Mid) MT: Vaynor IR⁵ Apr 2020Acer platanoides SN9152 Celsau CFA11: Treflys JC¹ Mar 2020 Acer platanoides SO2314 Llanelly Mons: Llanelly JC¹ Feb 2019Acer platanoides SN9758 Cwm Crogau CFA11: Llanafanfawr DB¹ Oct 2018 Acer platanoides SO0924 Castle Farm CFA12: Talybont-On-Usk DB¹ Jan 2018 Acer platanoides SN9208 Afon Mellte CFA15: Ystradfellte: SSSI0451, DB⁴ SAC71, IPA139 Apr 2017Acer platanoides SN9665 Wernnewydd CFA09: Llanwrthwl DB¹ Jul 2016 Acer platanoides SO0627 Usk CFA12: Llanfrynach DB¹ Jun 2015Acer platanoides SN8411 Coelbren CFA15: Tawe-Uchaf DB² Sep 2014Acer platanoides SO1937 Tregoyd Villa field CFA13: Gwernyfed DB¹ Jan 2014 Acer platanoides SO2316 Cwrt y Gollen site CFA14: Grwyney… DB¹ Apr 2012 Acer platanoides SO0528 Brecon CFA12: Brecon DB¹⁷ 2008 Acer platanoides SO1223 Llansantffraed CFA12: Talybont-On-Usk DB² May 2002Acer platanoides SO1940 Below Little Ffordd-fawr CFA13: Llanigon DB² Apr 2002Acer platanoides SO2142 Hay on Wye CFA13: Llanigon DB² Jul 2000 Acer platanoides SO2821 Pont -
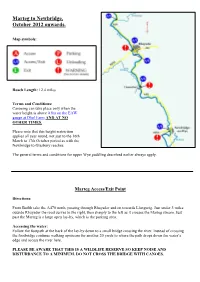
Marteg to Newbridge, October 2012 Onwards
Marteg to Newbridge, October 2012 onwards. Map symbols: Reach Length: 12.4 miles Terms and Conditions: Canoeing can take place only when the water height is above 0.8m on the EAW gauge at Dhol Farm AND AT NO OTHER TIMES. Please note that this height restriction applies all year round, not just to the 16th March to 17th October period as with the Newbridge to Glasbury reaches. The general terms and conditions for upper Wye paddling described earlier always apply. Marteg Access/Exit Point Directions: From Builth take the A470 north, passing through Rhayader and on towards Llangurig. Just under 3 miles outside Rhayader the road curves to the right, then sharply to the left as it crosses the Marteg stream. Just past the Marteg is a large open lay-by, which is the parking area. Accessing the water: Follow the footpath at the back of the lay-by down to a small bridge crossing the river. Instead of crossing the footbridge continue walking upstream for another 20 yards to where the path drops down the water’s edge and access the river here. PLEASE BE AWARE THAT THIS IS A WILDLIFE RESERVE SO KEEP NOISE AND DISTURBANCE TO A MINIMUM. DO NOT CROSS THE BRIDGE WITH CANOES. Rhayader Access/Exit Point (1) This exit point is for those who wish to avoid the grade IV rapids at Rhayader Bridge (highlighted by the exclamation mark on the map below), which can be a serious challenge to all but the most experienced paddler, especially in high water. Directions: From the centre of Rhayader (monument) take the B4518 (Bridge Street) in a westerly direction towards Elan Valley. -

Powys County Council Joint Housing Land Availability Study (2016)
Powys County Council Joint Housing Land Availability Study (2016) Between Powys County Council Local Planning Authority and the Study Group: Natural Resources Wales Barratt Homes Mid Wales Housing Hughes Architects Dwr Cymru Garner Southall Berrys Home Builders Federation Outdoor Recreation PCC McCartneys Melin Homes AB Planning Anne Price Mike Davies Nigel Griffiths 19 August 2016 Contents 1. Summary 2. Housing Land Supply Appendix 1 – Site Schedules Appendix 2 – Past Completions Data Appendix 3 – Previous Land Supply Data 1. Summary 1.1 This is the Powys County Council Land Availability Study (JHLAS) for 2016 which presents the housing land supply for the area at the base date of 1st April 2016. It replaces the report for the previous base date of 2015. 1.2 The JHLAS has been prepared in accordance with the requirements of Planning Policy Wales (PPW) and Technical Advice Note 1 (TAN 1). Please refer to these documents for details of the requirements for the maintenance of a five-year housing land supply in each local Planning Authority area and the process for undertaking the JHLASs. http://gov.wales/topics/planning/planningstats/housing-land-availability-in-wales/?lang=en http://gov.wales/topics/planning/planningstats/housing-land-availability-in-wales/?skip=1&lang=cy 1.3 Section 2 sets out details of the housing land supply and how it has been calculated. It shows that based on the Table 4 calculation method set out in TAN 1 Powys County Council has 2.2 years housing land supply. In regard to the calculation method used, TAN 1 states in situations where the adopted LDP only covers part of the 5 year study period, the average annual requirement form the LDP should be extrapolated to give an estimate of the land required (See Table 4 in Annex 3 for the formula for the calculation). -

SOUTH WALES. Fhayadert
DIRECTORY.] SOUTH WALES. FHAYADERt postmaster. Letters arrive from ~ part8 at 6.40 School At.te.ndanoe Oom.mittee. 1 a.m. & 5.20 p.m.; delivery commences at 1.15 8.m. Meets at Justices' rQOIWl on,. e'V~~ a~tet-nate w~. ~fJ:! p.m &; 5.30 'p.m, town OILY; &; dispatched at 6.50 p.m. to Clerk, George l\:lorgan Jarman, North street, Rhayadet all parts i box closing at 6.30 p.tu. & with extra !d. Attendance Officers" Richard Price, North at. 'Rhayad~ stamp at 6.35 p.m. Box cleared on sundays -at 6.30 &; John Griffiths. inn. Llandrindod p.m.; delivery commences at 9.40 ".m County Magistrates for the Petty Sessional Divudon of Rural Ilistrid Council. .. Rhayader. Meets at Justices' rOOmil on every alternate wedJ a' EvaUS' Edward Middleton esq. B.A. Llynbarried, Rhaya- 12 nOOn. der, chairman Clerk, Georg& Morgan Jarman, North stre~ Rhayader Carter John Corrie esq. Cefnfaes, Nantmel Treasurer, Geo. R8&, North &i South Wales Bank,Rhayadr Lewis-Uoyd Robert esq. D.L. Bryntirion, Nantmel Medical Officers of Health, Alfred GordOIi Richardson Lloyd-Verney Col. George Hope, Clochfaen, Llanidloes M.B., C.M. G~anant, Rhayader &; William Bowen Da.viel Morgan Richard esq. Llwyndre, Rhayader L.R.C.P.Lond. Llandrindod Wells Powell David Price esq. Howey hall, Llandrindod Sanitary Inspectors, Jamall Richard' Powell, Llanyre, Prickard Rev. Wm. Edward M.A. Dderw, Cwmdauddwr Llandrindod Wells R.S.O.; &; Robt. W.orthing,Rha.yadr Sladen Major-General, John Ramsey R.A., D.L. Rhydol- dog, Rhayader Publio Establishment~ Williams Edward esq. -

Newsletter Ebrill / April 2020 Rhif / No 139
The picture can't be displayed. Cymdeithas Melinau Cymru Welsh Mills Society NEWSLETTER EBRILL / APRIL 2020 RHIF / NO 139 Former windmill tower on Caldey Island (see pages 13-18) Cymdeithas Melinau Cymru Welsh Mills Society NEWSLETTER 139 April 2020 Contents: Editorial 3 The mills of Caldly Island 13 Dates for your Diary 4 Mills for Sale 19 Membership News 5 Twenty-five Years Ago 22 News from the mills 7 The Welsh Mills Society was launched in 1984. The aims of the Society are to study, record, interpret and publicise the wind and water mills of Wales, to encourage general interest, and to advise on their preservation and use. Officers and Committee Officers: Chairman: Gerallt Nash [email protected] Secretary: Hilary Malaws [email protected] Treasurer: Tim Haines [email protected] Membership Secretary: Tim Haines [email protected] Journal Editor: Mel Walters [email protected] (Coed Trewernau Mill, Crossgates, Llandindrod Wells, Powys, LD1 6PG) Committee: Gareth Beech [email protected] John Crompton (Mucky Mills) [email protected] Andrew Findon (Mill Owners’ Forum) [email protected] 01974 251231 Emma Hall [email protected] Anne Parry [email protected] Jane Roberts (Bring & Buy stall) 01633 780247 Helen Williams [email protected] For further information, please write to the Hon. Secretary: Hilary Malaws, Y Felin, Tynygraig, Ystrad Meurig, Ceredigion, Wales SY25 6AE or visit our website at: www.welshmills.org 2 GAIR GAN Y GOLYGYDD / EDITORIAL Mae’r misoedd diwethaf wedi bod yn rhai pryderus i lawer. Codwyd dŵr yr afonydd i lefelau dychrynllyd ar draws Cymru a Lloegr o ganlyniad i gyfnod hir o law trwm. -

Upper Wye Catchment Management Plan Consultation Report
N SLA- Ij/S 5 2 UPPER WYE CATCHMENT MANAGEMENT PLAN CONSULTATION REPORT N.R.A - Welsh Region REGIONAL TECHNICAL (PLANNING) Reference No : RTP017 LIBRARY COPY - DO NOT REMOVE RECYCLED PAPER A)£A V\I^GS 52- n a t io n a l RIVERS AUTHORITY . .WELSH REGION ____ - - - UPPER WYE CATCHMENT MANAGEMENT PLAN CONSULTATION REPORT National Rivers Authority - Welsh Region South East Area Rivers House St Mellons Business Park St Mellons Cardiff CF3 OLT June 1993 UPPER WYE CATCHMENT MANAGEMENT PLAN CONSULTATION REPORT CONTENTS PAGE No. FOREWORD iv MISSION STATEMENT OF THE NRA v THE NATIONAL RIVERS AUTHORITY vi 1.0 CONCEPT OF THE CATCHMENT MANAGEMENT PLAN 1 2.0 THE UPPER WYE CATCHMENT 4 2.1 Catchment Description 5 2.2 Data collection Within the Catchment 7 2.3 Key Details 8 3.0 CATCHMENT USES 9 3.1 Introduction 10 DEVELOPMENT AND LAND USE 3.2 Development 11 3.3 Flood Defence - 14 3.4 Forestry 17 3.5 Farming 19 CONSERVATION AND FISHERIES 3.6 Conservation - Ecology 20 3.7 Conservation - Landscape and Archaeology 24 3.8 Fisheries Ecosystem 26 3.9 Angling and Commercial Fishing 29 ABSTRACTIONS 3.10 Abstraction for Potable Water Supply - Groundwater 31 3.11 Abstraction for Potable Water Supply - Surface Water 34 3.12 Agricultural Abstraction 37 3.13 Livestock Watering 40 3.14 Industrial and Commercial Abstraction 41 3.15 Water Power 43 DISCHARGES AND POLLUTION CONTROL 3.16 Sewage and Trade Discharges 45 3.17 Waste Disposal to Land 47 AMENITY, NAVIGATION AND WATER SPORTS 3.18 Amenity 48 3.19 Navigation and Boating 50 3.20 Immersion Sports 52 4.0 CATCHMENT TARGETS 53 4T Introduction. -

Nov – Feb 2019 JOINT WALKS PROGRAMME (Incl
FOUR WELLS GROUP/EAST RADNOR GROUP GRWP Y FFYNHONNAU/GRWP DWYRAIN MAESYFED Web Site: fourwells.powysramblers.org.uk powysramblers.org.uk/east -radnor-group Nov – Feb 2019 JOINT WALKS PROGRAMME (incl. Four Wells Short Monday & Elan Valley Trust Walks) Date Description Grade Start / Map Ref. Leader November 8 miles Alan 3rd Llanwrthwl, Penrhos, Clyn, Carn Gafallt, Cefn Start at Llanwrthwl Village Hall SN974 638 10.00 am Energetic 01597 811051 Car share Llanelwedd Ent Park SO005517 at 9.15am 10 miles Wendy #10th Cwmbach, Ffynnon Gynydd, The Begwyns Start at OL13 SO 167396. Small lay-by on right next Energetic 01568 612556 to footbridge Meet at Llandrindod Wells railway station at 9.15 Builth Wells to Llandrindod Wells linear 10.5 miles Peter 17th for 9.25 bus. Part of the new Heart of Wales Line Trail Moderate 01982 570482 Start at 10.00am Heart of Wales Line Trail *Car share South of Crossgates opp. church 10 miles Kevin and Laura Llanbister Road to Knucklas SO087643 at 9.00 am #24th Moderate 01547 520266 LINEAR Exp201 SO252741 Park at Community Centre for 400m ascent 07975 876383 Knucklas Station to catch 9.59 train Richard 26 th Llanwrthwl & Dulas valley 5 miles easy Start at Llanwrthwl SN976638 at 10.00am 01597 810604 Monday December 9 miles 1st Crickadarn, Waun Gynllwch, Twmpath Start at Erwood Craft Centre SO089439 10am Rob moderate 01982 560471 *Car share South of Crossgates opp. church Jo 8.1 miles SO087643 at 9.15 am #8 th Thorn, Offa’s Dyke Path, Ackhill 07498 Moderate Start at Exp 201 SO 315642 Presteigne Recycling Car 489028 Park *Car share Llanelwedd Ent park SO 055 517 at 8 miles Chris P 15th Cwmbach (Glasbury) - The Begwns - Ffynnon Gynydd moderate 9.15am 01874 938397 Start SO167397 at 10am. -

Builth Wells to Glasbury
Builth Wells – Glasbury, December 2018 onwards Reach Length: 24.2 miles Terms and Conditions: Canoeing can take place between 18th October and 15th March inclusive. In the event of high water outside this period, additional access is granted when the water height on the gauge at Erwood http://www.wyeuskfoundation.org/conditions/gauge1.php is above the red marker. The general terms and conditions for upper Wye paddling described earlier always apply. Builth Wells Access Point (map on p3) Directions: Follow the Builth one-way system through the high street and round until you come to the Groe public car park on your left (entrance next to the Tourist Information Centre). Unloading: The car park is a short distance to the egress point on the right bank above the bridge. Bachawy Brook Access/Exit Point (map on p3) Directions, Unloading & Parking: From Builth Wells take the A470 south towards Brecon. After approximately 6 miles, turn left crossing the Wye over the Erwood Bridge (signposted to Aberedw and Painscastle). The road passes underneath another bridge before curving to the left. Take the next left (signposted to Boughrood & Llanstephan). After 1.3 miles the road crosses the Bachawy Brook via an old railway bridge. Immediately before this bridge there is a lay-by on your left. Park here and unload. Accessing the water: Using the path from the lay-by, walk down to the Bachawy Brook, crossing it via the small footbridge. Please note that this is a permissive path, not a right of way. On no account paddle down the Bachawy – there is a large watergate blocking your passage. -

Notice of Election Powys County Council - Election of Community Councillors
NOTICE OF ELECTION POWYS COUNTY COUNCIL - ELECTION OF COMMUNITY COUNCILLORS An election is to be held of Community Councillors for the whole of the County of Powys. Nomination papers must be delivered to the Returning Officer, County Hall, Llandrindod Wells, LD1 5LG on any week day after the date of this notice, but not later than 4.00pm, 4 APRIL 2017. Forms of nomination may be obtained at the address given below from the undersigned, who will, at the request of any elector for the said Electoral Division, prepare a nomination paper for signature. If the election is contested, the poll will take place on THURSDAY, 4 MAY 2017. Electors should take note that applications to vote by POST or requests to change or cancel an existing application must reach the Electoral Registration Officer at the address given below by 5.00pm on the 18 APRIL 2017. Applications to vote by PROXY must be made by 5.00pm on the 25 APRIL 2017. Applications to vote by PROXY on the grounds of physical incapacity or if your occupation, service or employment means you cannot go to a polling stations after the above deadlines must be made by 5.00 p.m. on POLLING DAY. Applications to be added to the Register of Electors in order to vote at this election must reach the Electoral Registration Officer by 13 April 2017. Applications can be made online at www.gov.uk/register-to-vote The address for obtaining and delivering nomination papers and for delivering applications for an absent vote is as follows: County Hall, Llandrindod Wells, LD1 5LG J R Patterson, Returning Officer -

St Gwrthwls Llanwrthwl to St Brides Cwmdauddwr
St. Gwrthwl’s, Llanwrthwl to St Brides, Cwmdauddwr (3 miles, mostly tarmac, fairly flat, one moderate hill) From the church turn left through the village, past the old school (now community centre) on the right and ignore the turning on the left to Penrhos. 50 yards further on, take the bridleway on the right up the hill. At the tarmac road, turn right. Follow the lane for 3/4 mile until you pass a house called Bach Y Graig on the left. A short distance after the road veers around to the left but follow a bridleway that leads straight on down to the river. Go through the gate and follow the path to the riverside. In the middle distance you can see a bridge over the Wye carrying the Mid Wales railway, closed in 1966. This is an ideal place for a paddle and if the river is low you can take off socks and shoes and wade to the other side. However, the proper footpath turns left and crosses the river by a rather scary looking suspension bridge. Despite the way it swings about a bit, this bridge is perfectly safe! Follow the footpath through a gate to Glyn Farm. Turn right onto the tarmac lane and follow the lane all the way to Cwmdauddwr past Cwmdauddwr Park on the right. Fork left at the Triangle Inn, a 16th Century drover’s Inn, and the church is on the left. Alternatively, from the farm follow the tarmac lane as far as a patch of woodland on the left. -

Traveline Cymru A4 Landscape Timetable
Llanidloes - Llandrindod Wells via Rhayader Service X47 (PWBX047) Monday to Friday (Excluding Bank Holidays) (Inbound) Timetable valid from 1st February 2013 until 26th May 2017 Llanidloes Gro 0635 0740 0950 -- 1355 -- 1640 Tylwch Old Station | | 1000 -- 1405 -- 1650 Nantgwyn | | 1005 -- 1410 -- 1655 Pant-y-dwr | | 1008 -- 1413 -- 1658 Saint Harmon Sun Inn | | 1012 -- 1417 -- 1702 Llangurig Black Lion 0643 0748 | 1206 | -- | Dolhelfa Ganol 0650 0755 | 1213 | -- | Rhayader Dark Lane TIC (arr) 0657 0802 1020 1220 1425 -- 1710 Rhayader Dark Lane TIC (dep) 0658 0808 1023 1223 1428 1638 1713 Llanwrthwl Turn 0703 0813 1028 1228 1433 1643 1718 Argoed Mill 0706 0816 1031 1231 1436 1646 1721 Newbridge-on-Wye Post Office | 0822 1037 1237 1442 1652 1727 Llanyre War Memorial | 0826 1041 1241 1446 1656 1731 Llandrindod Wells Brookland Road 0712 0830 1045 1245 1450 1700 1735 Llandrindod Wells Railway Station A 0715 0833 1048 1248 1453 1703 1738 Llandrindod Wells Powys County Hall -- 0836 -- -- -- -- -- Landrindod High School -- 0840 -- -- -- -- -- Llandrindod Wells - Llanidloes via Rhayader Service X47 (PWBX047) Monday to Friday (Excluding Bank Holidays) (Outbound) Timetable valid from 1st February 2013 until 26th May 2017 Notes: X Llandrindod High School -- -- -- -- -- 1603 -- -- Llandrindod Wells Powys County Hall -- -- -- -- -- 1607 1707 -- Llandrindod Wells Railway Station A 0717 -- 0910 1110 1305 1610 1710 1810 Llandrindod Wells Brookland Road 0720 -- 0913 1113 1308 1613 1713 1813 Llanyre War Memorial 0724 -- 0917 1117 1312 1617 1717 1817 Newbridge-on-Wye