PRIM1 Antibody Purified Rabbit Polyclonal Antibody (Pab) Catalog # AP51446
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Supplementary Text and Figures.Pdf
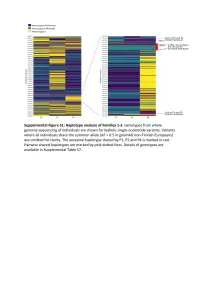
Supplemental Figure S2. PRIM1 immunoblots (20-75 kDa). (A) Corresponds to Figure 2D, with additional bands seen in P2 at 75 kDa and 25 kDa. (B) Independent experiment with cell lysates from C and P2 in which additional bands at 25 kDa not evident. (C) Corresponds to Figure 2D. No additional band visible at 25 kDa. (D) Validation of monoclonal antibody (8G10) with siRNA to PRIM1 in HeLa cells. siLUC, luciferase negative control. siPRIM1, siRNA targeting PRIM1 transcript. Vinculin used as a loading control. Supplemental Figure S3: Predicted destabilisation of PRIM1 by the C301R substitution. (A) Cysteine 301 lies in a buried hydrophobic region in PRIM1. DNA primase dimer crystal structure (PDB: 4BPU). PRIM1 residues shaded according to solvent accessibility. Cysteine 301, substituted to arginine in P5, lies in a buried hydrophobic region. (B) Cys301 is evolutionarily conserved in vertebrates. Neither arginine or other large or charged amino acids are observed at this position in other species. (C) The C301R substitution observed in P5 is predicted to lead to destabilisation of all available PRIM1 crystal structures. In contrast, substitution with leucine or threonine, as observed in some orthologous proteins, is not predicted to lead to destabilisation. Data points, predicted changes in free-energy (ΔΔG) from FoldX plotted for each of 14 available crystal structures. (D) Immunoblotting of RPE1 cells transfected with dual reporter constructs (as described in Figure 3B and Materials and Methods) confirms production of PRIM1-GFP and FLAG-SR at expected molecular weights and shows reduced protein levels for the C301R variant. n=2 experiments shown. C301R protein levels relative to WT after normalization to actin loading control indicated underneath the blot. -

The Cdc23 (Mcm10) Protein Is Required for the Phosphorylation of Minichromosome Maintenance Complex by the Dfp1-Hsk1 Kinase
The Cdc23 (Mcm10) protein is required for the phosphorylation of minichromosome maintenance complex by the Dfp1-Hsk1 kinase Joon-Kyu Lee*†, Yeon-Soo Seo†, and Jerard Hurwitz*‡ *Program in Molecular Biology, Memorial Sloan–Kettering Cancer Center, New York, NY 10021; and †Department of Biological Science, Korea Advanced Institute of Science and Technology, Daejeon 305-701, Korea Contributed by Jerard Hurwitz, December 4, 2002 Previous studies in Saccharomyces cerevisiae have defined an (9). Furthermore, studies in S. cerevisiae with mcm degron essential role for the Dbf4-Cdc7 kinase complex in the initiation of mutants showed that the Mcm proteins are also required for the DNA replication presumably by phosphorylation of target proteins, progression of the replication fork (10). These observations such as the minichromosome maintenance (Mcm) complex. We suggest that the Mcm complex is the replicative helicase. have examined the phosphorylation of the Mcm complex by the Cdc7, a serine͞threonine kinase conserved from yeast to Dfp1-Hsk1 kinase, the Schizosaccharomyces pombe homologue of humans (reviewed in ref. 11), is activated by the regulatory Dbf4-Cdc7. In vitro, the purified Dfp1-Hsk1 kinase efficiently phos- protein Dbf4. Although the level of Cdc7p is constant through- ͞ phorylated Mcm2p. In contrast, Mcm2p, present in the six-subunit out the cell cycle, the activity of this kinase peaks at the G1 S Mcm complex, was a poor substrate of this kinase and required transition, concomitant with the cellular level of Dbf4p. In S. ͞ Cdc23p (homologue of Mcm10p) for efficient phosphorylation. In cerevisiae, Dbf4p binds to chromatin at the G1 S transition and the presence of Cdc23p, Dfp1-Hsk1 phosphorylated the Mcm2p and remains attached to chromatin during S phase (12). -

Assembly of the Cdc45-Mcm2–7-GINS Complex in Human Cells Requires the Ctf4/And-1, Recql4, and Mcm10 Proteins
Assembly of the Cdc45-Mcm2–7-GINS complex in human cells requires the Ctf4/And-1, RecQL4, and Mcm10 proteins Jun-Sub Ima,1, Sang-Hee Kia,1, Andrea Farinab, Dong-Soo Junga, Jerard Hurwitzb,2, and Joon-Kyu Leea,2 aDepartment of Biology Education, Seoul National University, Seoul, 151-748, Korea; and bProgram in Molecular Biology, Memorial Sloan–Kettering Cancer Center, New York, NY 10021 Contributed by Jerard Hurwitz, July 17, 2009 (sent for review July 1, 2009) In eukaryotes, the activation of the prereplicative complex and Cdc45, and GINS (the CMG complex), purified from Drosophila assembly of an active DNA unwinding complex are critical but embryos, has DNA helicase activity in vitro (12). poorly understood steps required for the initiation of DNA repli- In addition to the above components, the assembly of the DNA cation. In this report, we have used bimolecular fluorescence unwinding complex also depends on additional factors including complementation assays in HeLa cells to examine the interactions Dpb11/Cut5, Sld2, Sld3, and Mcm10. Mcm10 is required for the between Cdc45, Mcm2–7, and the GINS complex (collectively called chromatin binding of Cdc45 and DNA polymerase ␣ in yeast and the CMG complex), which seem to play a key role in the formation Xenopus (13–16). Dpb11/Cut5 is essential for origin binding of and progression of replication forks. Interactions between the GINS in Saccharomyces cerevisiae (17), and reported to be required CMG components were observed only after the G1/S transition of for chromatin binding of Cdc45 and DNA polymerase ␣ in Xenopus the cell cycle and were abolished by treatment of cells with either (18). -

Mcm10 Has Potent Strand-Annealing Activity and Limits Translocase-Mediated Fork Regression
Mcm10 has potent strand-annealing activity and limits translocase-mediated fork regression Ryan Maylea, Lance Langstona,b, Kelly R. Molloyc, Dan Zhanga, Brian T. Chaitc,1,2, and Michael E. O’Donnella,b,1,2 aLaboratory of DNA Replication, The Rockefeller University, New York, NY 10065; bHoward Hughes Medical Institute, The Rockefeller University, New York, NY 10065; and cLaboratory of Mass Spectrometry and Gaseous Ion Chemistry, The Rockefeller University, New York, NY 10065 Contributed by Michael E. O’Donnell, November 19, 2018 (sent for review November 8, 2018; reviewed by Zvi Kelman and R. Stephen Lloyd) The 11-subunit eukaryotic replicative helicase CMG (Cdc45, Mcm2-7, of function using genetics, cell biology, and cell extracts have GINS) tightly binds Mcm10, an essential replication protein in all identified Mcm10 functions in replisome stability, fork progres- eukaryotes. Here we show that Mcm10 has a potent strand- sion, and DNA repair (21–25). Despite significant advances in the annealing activity both alone and in complex with CMG. CMG- understanding of Mcm10’s functions, mechanistic in vitro studies Mcm10 unwinds and then reanneals single strands soon after they of Mcm10 in replisome and repair reactions are lacking. have been unwound in vitro. Given the DNA damage and replisome The present study demonstrates that Mcm10 on its own rap- instability associated with loss of Mcm10 function, we examined the idly anneals cDNA strands even in the presence of the single- effect of Mcm10 on fork regression. Fork regression requires the strand (ss) DNA-binding protein RPA, a property previously unwinding and pairing of newly synthesized strands, performed by associated with the recombination protein Rad52 (26). -

Gene Expression Profiling Analysis Contributes to Understanding the Association Between Non-Syndromic Cleft Lip and Palate, and Cancer
2110 MOLECULAR MEDICINE REPORTS 13: 2110-2116, 2016 Gene expression profiling analysis contributes to understanding the association between non-syndromic cleft lip and palate, and cancer HONGYI WANG, TAO QIU, JIE SHI, JIULONG LIANG, YANG WANG, LIANGLIANG QUAN, YU ZHANG, QIAN ZHANG and KAI TAO Department of Plastic Surgery, General Hospital of Shenyang Military Area Command, PLA, Shenyang, Liaoning 110016, P.R. China Received March 10, 2015; Accepted December 18, 2015 DOI: 10.3892/mmr.2016.4802 Abstract. The present study aimed to investigate the for NSCL/P were implicated predominantly in the TGF-β molecular mechanisms underlying non-syndromic cleft lip, signaling pathway, the cell cycle and in viral carcinogenesis. with or without cleft palate (NSCL/P), and the association The TP53, CDK1, SMAD3, PIK3R1 and CASP3 genes were between this disease and cancer. The GSE42589 data set found to be associated, not only with NSCL/P, but also with was downloaded from the Gene Expression Omnibus data- cancer. These results may contribute to a better understanding base, and contained seven dental pulp stem cell samples of the molecular mechanisms of NSCL/P. from children with NSCL/P in the exfoliation period, and six controls. Differentially expressed genes (DEGs) were Introduction screened using the RankProd method, and their potential functions were revealed by pathway enrichment analysis and Non-syndromic cleft lip, with or without cleft palate (NSCL/P) construction of a pathway interaction network. Subsequently, is one of the most common types of congenital defect and cancer genes were obtained from six cancer databases, and affects 3.4-22.9/10,000 individuals worldwide (1). -

PRIM1 Polyclonal ANTIBODY Catalog Number:10773-1-AP 2 Publications
For Research Use Only PRIM1 Polyclonal ANTIBODY www.ptglab.com Catalog Number:10773-1-AP 2 Publications Catalog Number: GenBank Accession Number: Purification Method: Basic Information 10773-1-AP BC005266 Antigen affinity purification Size: GeneID (NCBI): Recommended Dilutions: 150UL , Concentration: 133 μg/ml by 5557 WB 1:500-1:1000 Bradford method using BSA as the Full Name: IP 0.5-4.0 ug for IP and 1:200-1:1000 standard; primase, DNA, polypeptide 1 (49kDa) for WB Source: IHC 1:20-1:200 Calculated MW: IF 1:50-1:500 Rabbit 50 kDa Isotype: Observed MW: IgG 50 kDa Immunogen Catalog Number: AG1124 Applications Tested Applications: Positive Controls: IF, IHC, IP, WB, ELISA WB : K-562 cells, HeLa cells, Sp2/0 cells Cited Applications: IP : HeLa cells, IHC IHC : human lymphoma tissue, Species Specificity: human, mouse, rat IF : HeLa cells, Cited Species: mouse Note-IHC: suggested antigen retrieval with TE buffer pH 9.0; (*) Alternatively, antigen retrieval may be performed with citrate buffer pH 6.0 DNA replication in human cells is initiated by a complex apparatus containing a DNA polymerase-alpha/primase Background Information complex. Primase synthesizes oligoribonucleotides that serve as primers for the initiation of DNA synthesis. It plays a role in both the initiation of DNA replication and the synthesis of Okazaki fragments for lagging strand synthesis. PRIM1 is a subnuit of this complex. Notable Publications Author Pubmed ID Journal Application Pierre Kunz 30986223 PLoS One IHC Liam C Hunt 31365869 Cell Rep Storage: Storage Store at -20°C. Stable for one year after shipment. -

Mcm10 and And-1/CTF4 Recruit DNA Polymerase to Chromatin For
Downloaded from genesdev.cshlp.org on September 26, 2021 - Published by Cold Spring Harbor Laboratory Press Mcm10 and And-1/CTF4 recruit DNA polymerase ␣ to chromatin for initiation of DNA replication Wenge Zhu,1,3 Chinweike Ukomadu,1,3 Sudhakar Jha,1,3 Takeshi Senga,1 Suman K. Dhar,1 James A. Wohlschlegel,1 Leta K. Nutt,2 Sally Kornbluth,2 and Anindya Dutta1,4 1Department of Biochemistry and Molecular Genetics, University of Virginia School of Medicine, Charlottesville, Virginia 22908, USA; 2Department of Pharmacology and Cancer Biology, Duke University Medical Center, Durham, North Carolina 27710, USA The MCM2-7 helicase complex is loaded on DNA replication origins during the G1 phase of the cell cycle to license the origins for replication in S phase. How the initiator primase–polymerase complex, DNA polymerase ␣ (pol ␣), is brought to the origins is still unclear. We show that And-1/Ctf4 (Chromosome transmission fidelity 4) interacts with Mcm10, which associates with MCM2-7, and with the p180 subunit of DNA pol ␣. And-1 is essential for DNA synthesis and the stability of p180 in mammalian cells. In Xenopus egg extracts And-1 is loaded on the chromatin after Mcm10, concurrently with DNA pol ␣, and is required for efficient DNA synthesis. Mcm10 is required for chromatin loading of And-1 and an antibody that disrupts the Mcm10–And-1 interaction interferes with the loading of And-1 and of pol ␣, inhibiting DNA synthesis. And-1/Ctf4 is therefore a new replication initiation factor that brings together the MCM2-7 helicase and the DNA pol ␣–primase complex, analogous to the linker between helicase and primase or helicase and polymerase that is seen in the bacterial replication machinery. -

Single-Molecule Visualization of Saccharomyces Cerevisiae Leading-Strand Synthesis Reveals Dynamic Interaction Between MTC and the Replisome
Single-molecule visualization of Saccharomyces cerevisiae leading-strand synthesis reveals dynamic interaction between MTC and the replisome Jacob S. Lewisa,b,1, Lisanne M. Spenkelinka,b,c,1, Grant D. Schauerd, Flynn R. Hilla,b, Roxanna E. Georgescud, Michael E. O’Donnelld,2, and Antoine M. van Oijena,b,2 aCentre for Medical & Molecular Bioscience, University of Wollongong, Wollongong, NSW 2522, Australia; bIllawarra Health & Medical Research Institute, Wollongong, NSW 2522, Australia; cZernike Institute for Advanced Materials, University of Groningen, 9747 AG Groningen, The Netherlands; and dHoward Hughes Medical Institute, Rockefeller University, New York, NY 10065 Contributed by Michael E. O’Donnell, August 24, 2017 (sent for review June 23, 2017; reviewed by David Rueda and Michael Trakselis) The replisome, the multiprotein system responsible for genome fork, additional proteins are conscripted to the complex to form duplication, is a highly dynamic complex displaying a large number the RPC. These proteins include Ctf4, Csm3, FACT, Mrc1, Pol of different enzyme activities. Recently, the Saccharomyces cerevisiae α, Tof1, and Top1 (8). It has been shown that Mrc1, a yeast minimal replication reaction has been successfully reconstituted in homolog of Claspin and an S-phase-specific mediator protein of vitro. This provided an opportunity to uncover the enzymatic activities the DNA damage response, is recruited to the fork (8, 9) and of many of the components in a eukaryotic system. Their dynamic increases the rate of replication in vivo about twofold (10–12). In behavior and interactions in the context of the replisome, however, vitro studies confirm that Mrc1 increases the speed of replication remain unclear. -

Consequences and Resolution of Transcription–Replication Conflicts
life Review Consequences and Resolution of Transcription–Replication Conflicts Maxime Lalonde †, Manuel Trauner †, Marcel Werner † and Stephan Hamperl * Institute of Epigenetics and Stem Cells (IES), Helmholtz Zentrum München, 81377 Munich, Germany; [email protected] (M.L.); [email protected] (M.T.); [email protected] (M.W.) * Correspondence: [email protected] † These authors contributed equally. Abstract: Transcription–replication conflicts occur when the two critical cellular machineries respon- sible for gene expression and genome duplication collide with each other on the same genomic location. Although both prokaryotic and eukaryotic cells have evolved multiple mechanisms to coordinate these processes on individual chromosomes, it is now clear that conflicts can arise due to aberrant transcription regulation and premature proliferation, leading to DNA replication stress and genomic instability. As both are considered hallmarks of aging and human diseases such as cancer, understanding the cellular consequences of conflicts is of paramount importance. In this article, we summarize our current knowledge on where and when collisions occur and how these en- counters affect the genome and chromatin landscape of cells. Finally, we conclude with the different cellular pathways and multiple mechanisms that cells have put in place at conflict sites to ensure the resolution of conflicts and accurate genome duplication. Citation: Lalonde, M.; Trauner, M.; Keywords: transcription–replication conflicts; genomic instability; R-loops; torsional stress; common Werner, M.; Hamperl, S. fragile sites; early replicating fragile sites; replication stress; chromatin; fork reversal; MIDAS; G-MiDS Consequences and Resolution of Transcription–Replication Conflicts. Life 2021, 11, 637. https://doi.org/ 10.3390/life11070637 1. -

R‐Loop Formation During S Phase Is Restricted by Primpol‐Mediated
Published online: November 26, 2018 Article R-loop formation during S phase is restricted by PrimPol-mediated repriming Sasa Svikovic1, Alastair Crisp1, Sue Mei Tan-Wong2, Thomas A Guilliam3, Aidan J Doherty3, Nicholas J Proudfoot2, Guillaume Guilbaud1 & Julian E Sale1,* Abstract sequences pose a barrier to DNA synthesis in vivo and the conse- quences of their doing so are not well understood. During DNA replication, conflicts with ongoing transcription are It is well established that long repetitive tracts lead to problems frequent and require careful management to avoid genetic insta- with both replication and transcription. For example, a long tract of bility. R-loops, three-stranded nucleic acid structures comprising a polypurine–polypyrimidine (GAA)n repeats (in which n can exceed DNA:RNA hybrid and displaced single-stranded DNA, are important 1,500) is linked to the inherited neurodegenerative disorder Friedre- drivers of damage arising from such conflicts. How R-loops stall ich’s ataxia (Campuzano et al, 1996). These repeats can form H- replication and the mechanisms that restrain their formation DNA (Frank-Kamenetskii & Mirkin, 1995), a triplex DNA structure during S phase are incompletely understood. Here, we show in vivo able to block replication both in bacterial, yeast and human cells how R-loop formation drives a short purine-rich repeat, (GAA)10,to (Ohshima et al, 1998; Krasilnikova & Mirkin, 2004; Chandok et al, become a replication impediment that engages the repriming 2012), which can promote genetic instability of the repeat (Gerhardt activity of the primase-polymerase PrimPol. Further, the absence et al, 2016). Furthermore, these repetitive tracts are prone to form of PrimPol leads to significantly increased R-loop formation around R-loops (Groh et al, 2014), three-stranded nucleic acid structures in this repeat during S phase. -

Control of Eukaryotic DNA Replication Initiation—Mechanisms to Ensure Smooth Transitions
G C A T T A C G G C A T genes Review Control of Eukaryotic DNA Replication Initiation—Mechanisms to Ensure Smooth Transitions Karl-Uwe Reusswig and Boris Pfander * Max Planck Institute of Biochemistry, DNA Replication and Genome Integrity, 82152 Martinsried, Germany; [email protected] * Correspondence: [email protected] Received: 31 December 2018; Accepted: 25 January 2019; Published: 29 January 2019 Abstract: DNA replication differs from most other processes in biology in that any error will irreversibly change the nature of the cellular progeny. DNA replication initiation, therefore, is exquisitely controlled. Deregulation of this control can result in over-replication characterized by repeated initiation events at the same replication origin. Over-replication induces DNA damage and causes genomic instability. The principal mechanism counteracting over-replication in eukaryotes is a division of replication initiation into two steps—licensing and firing—which are temporally separated and occur at distinct cell cycle phases. Here, we review this temporal replication control with a specific focus on mechanisms ensuring the faultless transition between licensing and firing phases. Keywords: DNA replication; DNA replication initiation; cell cycle; post-translational protein modification; protein degradation; cell cycle transitions 1. Introduction DNA replication control occurs with exceptional accuracy to keep genetic information stable over as many as 1016 cell divisions (estimations based on [1]) during, for example, an average human lifespan. A fundamental part of the DNA replication control system is dedicated to ensure that the genome is replicated exactly once per cell cycle. If this control falters, deregulated replication initiation occurs, which leads to parts of the genome becoming replicated more than once per cell cycle (reviewed in [2–4]). -

In Utero Exposure to Chlordecone Affects Histone Modifications And
In utero exposure to chlordecone affects histone modifications and activates LINE-1 in cord blood Louis Legoff, Shereen Cynthia D’cruz, Katia Bouchekhchoukha, Christine Monfort, Christian Jaulin, Luc Multigner, Fatima Smagulova To cite this version: Louis Legoff, Shereen Cynthia D’cruz, Katia Bouchekhchoukha, Christine Monfort, Christian Jaulin, et al.. In utero exposure to chlordecone affects histone modifications and activates LINE- 1 in cord blood. Life Science Alliance, Life Science Alliance LLC, 2021, 4 (6), pp.e202000944. 10.26508/lsa.202000944. hal-03217292 HAL Id: hal-03217292 https://hal.archives-ouvertes.fr/hal-03217292 Submitted on 4 May 2021 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Distributed under a Creative Commons Attribution| 4.0 International License Published Online: 9 April, 2021 | Supp Info: http://doi.org/10.26508/lsa.202000944 Downloaded from life-science-alliance.org on 4 May, 2021 Research Article In utero exposure to chlordecone affects histone modifications and activates LINE-1 in cord blood Louis Legoff1, Shereen Cynthia D’Cruz1, Katia Bouchekhchoukha1, Christine Monfort1, Christian Jaulin2, Luc Multigner1 , Fatima Smagulova1 Environmental factors can induce detrimental consequences into Epigenetic reprogramming during development is associated with a adulthood life.