Phytoplankton Assemblages in Two Sub-Basins of Lake Como
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
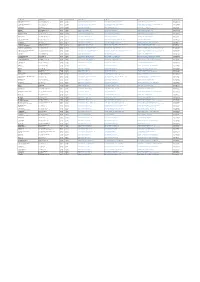
Comune Indirizzo Cap Codice Istat Sito Web E-Mail Pec
COMUNE INDIRIZZO CAP CODICE ISTAT SITO WEB E-MAIL PEC TELEFONO ABBADIA LARIANA Via Nazionale ,120 23821 97001 www.comune.abbadia-lariana.lc.it [email protected] [email protected] 0341731241 AIRUNO Via Pizzagalli Magno, 4 23881 97002 www.comune.airuno.lc.it [email protected] [email protected] 0399943222 ANNONE DI BRIANZA Via S. Antonio, 4 23841 97003 www.comune.annone-di-brianza.lc.it [email protected] [email protected] 0341576063 BALLABIO Via Valgrande, 4 23811 97004 www.comune.ballabio.lc.it [email protected] [email protected] 0341530111 BARZAGO Via Roma, 2 23890 97005 www.comune.barzago.lc.it [email protected] [email protected] 031860227 BARZANO' Viale Luciano Manara ,13 23891 97006 www.comune.barzano.lc.it [email protected] [email protected] 039921301 BARZIO Via A. Manzoni, 12 23816 97007 www.comune.barzio.lc.it [email protected] [email protected] 0341996125 BELLANO Via V. Veneto, 23 23822 97008 www.comune.bellano.lc.it [email protected] [email protected] 0341821124 BOSISIO PARINI P.zza G. Parini, 1 23842 97009 www.comune.bosisioparini.lc.it [email protected] [email protected] 0313580511 BRIVIO Via Vittorio Emanuele II, 12 23883 97010 www.comune.brivio.lc.it [email protected] [email protected] 0395320114 BULCIAGO Via Dante Alighieri, 2 23892 97011 www.comune.bulciago.lc.it [email protected] [email protected] 031860121 CALCO Via Italia, 25 23885 97012 www.comune.calco.lc.it [email protected] [email protected] 0399910017 CALOLZIOCORTE P.zza V. -

Comune Sede Di Seggio Orario Sede Indirizzo
ELENCO SEDI PRIMARIE SEGRETARIO REGIONALE /CONGRESSI PROVINCIALE E DI CIRCOLO COMUNE SEDE DI SEGGIO ORARIO SEDE INDIRIZZO COMUNI AFFERENTI 1 ABBADIA LARIANA 9,00 -14,00 SALA CIVICA VIA NAZIONALE ABBADIA LARIANA , MANDELLO DEL LARIO ,LIERNA 2 BRIVIO 9,00 -14,00 SALA CIVICA VIA V.EMANUELE 22 BRIVIO / AIRUNO 3 BARZAGO 9,00 -14,00 SEDE PD P.ZZA GARIBALDI BARZAGO 4 BARZANO'' 9,00 -14,00 SEDE PD VIA SIRTORI BARZANO'/BULCIAGO /CASSAGO B., NIBIONNO,CREMELLA/SIRTORI/VIGANO' 5 BELLANO 9,00 -14,00 SALA CIVICA VIA MARINAI D'ITALIA BELLANO , DERVIO,PERLEDO,VALVARRONE,VARENNA 6 BOSISIO PARINI 9,00 -14,00 AULA CIVICA P.ZZA PARINI BOSISIO PARINI 7 CALCO 9,00 -14,00 SALA CIVICA VIA VOLTA CALCO 8 CALOLZIOCORTE 9,00 -14,00 SEDE P.D. P.ZZA V.VENETO 14 CALOLZIOCORTE, ERVE , VERCURAGO 9 CASATENOVO 9,00 -14,00 SEDE PD VIA VERDI CASATENOVO/MONTICELLO 10 COLICO 9,00 -14,00 SALA CONSILIARE P.ZZA V° ALPINI COLICO 11 COSTA MASNAGA 9,00 -14,00 BIBLIOTECA COMUNALE P.ZZA CONFALONIERI 4 COSTA MASNAGA/ROGENO 12 GALBIATE 9,00 -14,00 SEDE PD VIA VENETO 15 GALBIATE 13 IMBERSAGO 9,00 -14,00 CIRCOLO COOPERATIVO VIA MATTEI IMBERSAGO 14 LECCO 9,00 - 18,00 CIRCOLO LIBERO PENSIERO VIA CALLONI 16 LECCO, SEZ.18,19,20,21,22,23 - BALLABIO 15 LECCO 9,00 - 18,00 CIRCOLO F.LLI FIGINI VIA ARMONIA LECCO SEZ, 28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43 16 LECCO 9,00 - 18,00 CAMPER + GAZEBO P.ZZA GARIBALDI LECCO SEZ. -
Simple Ways from the Museums to the Territory ITINERARIES Simple Ways
FROM THE MUSEUMS TO THE TERRITORY ITINERARIES SIMPLe Ways FROM THE MUSEUMS TO THE TERRITORY ITINERARIES SIMPLe Ways Edited by The province of Lecco Culture Service, Tourism and Sport Network of the Museums of the province of Lecco Planning and Coordination Anna Ranzi in collaboration with Scientific and Technical Committee of the Network of the Museums Editing Eleonora Massai Graphic Design and Printing Cattaneo Paolo Grafiche s.r.l. Oggiono - Annone B.za March 2018 (IIIth Edition) 2 INTRODUCTION The 2018 edition of the cultural tourist itineraries “SIMPLe Ways from the museums to the territory” is only one of many initiatives to help visitors rediscover and enjoy the rich and varied cultural heritage of the province of Lecco. This publication aims to provide the visitor with interest- ing ways to discover the collections in the Lecco Museum System, which counts a total of 30 museums to date. The aim is also to lead the visitor to extend their visit to the area itself with all its heritage sites and multifaceted beauty so that it becomes the real museum to explore. We have created a virtuous network of itineraries which allow local or tourist to visit the area and enjoy the landscape and natural surroundings with an increased awareness of the historic, artistic and architectural heritage. SIMPLe Ways are ten tourist itineraries exploring the Lecco branch of Lake Como, Valsassina, Val San Martino and Brianza, worthwhile destinations for visitors to the area who want to immerse themselves in the spectacular natural surroundings which still bear traces of the local heritage, at times until recently forgotten and only now rebuilt or restored. -

Case Turistiche: Più Compravendite a Varenna E Abbadia | 1
Case turistiche: più compravendite a Varenna e Abbadia | 1 Una veduta di Abbadia Lariana ABBADIA / VARENNA / LECCO – Cresce, pur lievemente rispetto al 2015, il volume di compravendite per il mercato residenziale turistico degli immobili nel lecchese, in un contesto dove l’offerta si è lievemente ridotta. E’ quanto emerge nel report dell’Ufficio Studi Gabetti su dati Gabetti, Professionecasa e Grimaldi. Nel 2016 le località monitorate sul Lario sono state contrassegnate da dinamiche differenti a seconda delle specifiche zone. Il ramo comasco, complessivamente più positivo per quello che ha riguardato l’andamento delle transazioni, ha visto anche una ripresa della domanda turistica, grazie alla maggiore presenza di interessanti soluzioni in termini di rapporto qualità/prezzo. Per quanto riguarda il ramo di Lecco, si segnala una buona richiesta turistica per Varenna, Mandello del Lario, e Lierna. Sulla sponda lecchese si è complessivamente riscontrata una stabilità delle quotazioni ad eccezione di Lierna, dove si è rilevata una lieve flessione. Varenna I tempi medi di vendita si sono attestati intorno ai 6-7 mesi, mentre gli sconti in sede di chiusura hanno una invece media del 10-12%. Per quanto riguarda le compravendite, si riscontrano dinamiche differenti a seconda delle Documento creato dal sito lecconotizie.com Case turistiche: più compravendite a Varenna e Abbadia | 2 specifiche realtà considerate. Secondo i dati dell’Agenzia del Territorio, nel 2015 si è osservato un lieve calo del numero di transazioni a Lecco, che sono passate da 377 nel 2014 a 341 nel 2015. In calo anche Bellano, che è passato dalle 51 transazioni registrate nel 2014 alle 33 del 2015. -

Provincia Di Lecco
LLEECCCCOO EE IILL LLAAGGOO Il territorio del Lecchese si offre al visitatore vario e ricco di paesaggi differenti, che creano un’impressionante suggestione di colori e atmosfere. La provincia di Lecco si estende ad est del Lago di Como abbracciandone le acque; il lago oppone all’imponente cornice delle montagne un dolce spettacolo di splendide ville immerse in parchi secolari. La visita si svolge - rà pertanto seguendo vari fili d’attrazione che formano una trama multicolore e preziosa: la natura e la sua valorizzazione, gli insediamenti abitati che punteggiano il paesaggio, gli episodi architettonici con particolare riguardo al periodo roma - nico. La proposta è quella di itinerari tematici diversi che favoriscono una approfondita conoscenza attraverso un’indagine storico-artistica ma anche naturalistica ed archeologica. Paesaggi è il titolo che abbiamo voluto dare a questa serie coordinata di tre pubblicazioni, proprio per rispecchiare le iden - tità di un territorio composito, che si caratterizza per tre aree differenti: il lago, la montagna e la pianura. Il risultato è un territorio tutto da scoprire, attraverso gli itinerari e le immagini raccolte in questa nuova pubblicazione coordinata che la Provincia di Lecco ha realizzato sviluppando una serie di proposte e percorsi individuati quali vere e proprie eccellenze. Oggi sta diventando una tendenza diffusa, anzi un vero e proprio fenomeno turistico, sicuramente il più nuovo, lo sviluppo dei “piccoli viaggi” alla ricerca di un luogo da esplorare, un borgo da visitare, un monumento da conoscere; viaggi dotati di un fascino e di un gusto irresistibili. Il turista è ormai abituato ai circuiti classici, è incuriosito ed attratto dalle zone più di nic - chia; non si accontenta soltanto di visitare chiese e palazzi ma è alla ricerca di percorsi tematici, di un patrimonio cultura - le di grande richiamo, di un vero viaggio d’arte e di cultura che offra nuove e continue emozioni. -

I COMUNI Di ABBADIA LARIANA, BELLANO, BOSISIO PARINI
I COMUNI di ABBADIA LARIANA, BELLANO, BOSISIO PARINI, COLICO, DERVIO, ESINO LARIO LIERNA, MANDELLO DEL LARIO, MOGGIO, VARENNA E LA RETE DI IMPRESE MONTAGNE DEL LAGO DI COMO in collaborazione con gli uffici di Piano dell’Ambito di Lecco e Bellano propongono ESTATE NEGLI UFFICI TURISTICI E NEI MUSEI nell’ambito del progetto LIVING LAND L’iniziativa si rivolge a ragazzi di età compresa tra i 18 e i 27 anni che desiderano cimentarsi in un’esperienza di crescita personale e di utilità per la comunità COSA E’ POSSIBILE FARE? I ragazzi selezionati saranno impegnati nelle attività degli uffici turistici e/o dei musei di Abbadia Lariana, Bellano, Bosisio Parini, Colico, Dervio, Esino Lario, Lierna, Mandello del Lario, Moggio, Varenna, o nella sede della rete d’imprese Montagne del lago di Como nel periodo compreso tra giugno e settembre 2018. Le attività consisteranno in: - Assistenza turistica diretta/telefonica/via mail - Aggiornamento sito web e pagine facebook - Gestione contatti con strutture turistiche - Cura esposizione del materiale - Presidio siti museali, storici, turistici - Visite guidate (in seguito ad opportuna formazione) Nello specifico: ENTE OSPITANTE LUOGHI DI ATTIVITA’ ORARI DI PERIODO MONTE APERTURA (SU ORE TURNI) Ufficio turistico di Ufficio turistico; Civico Lun-ven 9.30- 2 giugno-2 15-30 ore Abbadia Lariana museo Setificio Monti; 12.30;14-17 settembre settimanali possibili visite guidate sul Sab-Dom 8-14 territorio Ufficio turistico di Ufficio turistico; Orrido Lun-dom 11 giugno-30 30 ore Bellano 9.30-13.30; 15- settembre -

Ambito Di Bellano
AMBITO DI BELLANO Comuni Area Distrettuale di Bellano Segreteria operativa c/o Comunità Montana Valsassina Valvarrone Val d’Esino e Riviera - “Villa Merlo” Via Fornace Merlo, 2 – 23816 Barzio Tel. 0341-910144 int. 1 Fax. 0341-911640 e-mail: [email protected] ELENCO ASSISTENTI SOCIALI SERVIZIO SOCIALE DI BASE AMBITO DI BELLANO Orario sportelli aperti al pubblico GIORNI E ORARIO N° COMUNE/POLO ASSISTENTE SOCIALE PRESIDIO C/O RECAPITO DI RICEVIMENTO Tel. 0341/731241 1 Abbadia Lariana Dr.ssa Alessandra Bianchi Comune Lunedì e Venerdì: 10.00/13.00 e-mail: [email protected] Lunedì e Mercoledì: 9.00/12.00 Tel. 0341/530111 2 Ballabio Dr.ssa Annalisa Mariani Comune Giovedì: 16.00/19.00 e-mail: [email protected] 2° - 4° Sabato del mese: 9.00/12.00 Polo Altopiano (Comuni Comunità Montana Valsassina di Barzio, Moggio, Martedì: 9.00/13.00 Tel. 0341/910144 int. 1 3 Dr.ssa Elena Bertelle Valvarrone Val d'Esino e Riviera - Cremeno, Cassina V., Giovedì: 13.30/17.30 e-mail: [email protected] Barzio Morterone) Martedì e Giovedì: 09.00/11.30 Tel. 0341/821124 int. 5 4 Bellano Dr.ssa Alessandra Bianchi Comune di Bellano e-mail: [email protected] Tel. 0341/840123 Polo Altavalle Comune di Casargo I e III lunedì del mese 9.00/11.00 e-mail: [email protected] (Comuni di Taceno, 5 Dr.ssa Cassanelli Elisabetta Tel. 0341/880112 Crandola, Margno, Comune di Taceno II e IV lunedì del mese 9.00/11.00 Premana, Casargo, e-mail: [email protected] Pagnona, Parlasco) Lunedì 15.00/17.00 Tel. -

Calendario Completo Degli Incontri
CENTRO SPORTIVO ITALIANO Campionato ZONA LAGHI CAL Comitato di : LECCO Disciplina: CALCIO Coordinamento Attività Sportiva Categoria: UNDER 10 A.S. 2018/19 Girone: Primaverile – GIRONE “B” CALENDARIO COMPLETO DEGLI INCONTRI Andata: 1° Giornata Data e orario Località 1 CS Abbadia Lariana Blu - Oratorio Cassago A sab 09 feb 19 16:15 Abbadia Lariana - Oratorio P. Frassati 2 Us Orobia Robbiate - Polisp. Rovinata Bianca sab 09 feb 19 15:00 Robbiate - Oratorio . 3 Polisp. Rovinata Rossa - Gso Lomagna 2010 sab 09 feb 19 11:00 Lecco - Germanedo - Oratorio . 4 Orat. Cassago B - Gso Paderno d'Adda sab 09 feb 19 14:00 Cassago - Oratorio . 5 Garlate-Pescate - PCG Valmadrera "A" sab 09 feb 19 14:30 Garlate - Oratorio . Andata: 2° Giornata Data e orario Località 6 Gso Paderno d'Adda - Garlate-Pescate dom 17 feb 19 14:30 Paderno d'Adda - Oratorio . 7 Gso Lomagna 2010 - Orat. Cassago B dom 17 feb 19 00:00 Lomagna - Oratorio . 8 Polisp. Rovinata Bianca - Polisp. Rovinata Rossa sab 16 feb 19 11:00 Lecco - Germanedo - Oratorio . 9 Oratorio Cassago A - Us Orobia Robbiate sab 16 feb 19 14:00 Cassago - Oratorio . 10 PCG Valmadrera "A" - CS Abbadia Lariana Blu sab 16 feb 19 14:00 Valmadrera – Oratorio Via Bovara 3° Prova Polisportiva: Sabato 23 e domenica 24 Febbraio 2019 – Giochi di pallamano e pallacanestro Andata: 3° Giornata Data e orario Località 11 Us Orobia Robbiate - CS Abbadia Lariana Blu sab 02 mar 19 15:00 Robbiate - Oratorio . 12 Polisp. Rovinata Rossa - Oratorio Cassago A sab 02 mar 19 11:00 Lecco - Germanedo - Oratorio . 13 Orat. -

Viandante Inglese Sulle Orme Del Viandante 1
COMPLETE ITINERARY History road direct to the Stelvio’s pass (built between 1817 and 1834 ) was The oriental side of the Lake of opened; this caused the Como has been interested since abandonment of the previous road antiquity by a series of path that system, now call ed “The connected Milan with Alpine Wayfarer’s Trail” . It was passes. rediscovered recently and enhanced by the Company for promoting tourism in the Province of Lecco. The path start s shortly before Abbadia Lariana , near St Martino’s church. The path continues along the lakeshore, and then it reaches Historical postcard: the church of S t George in Varenna view from Fiumelatte Mandello del Lario , and continues to Maggiana, Rongio and Sonvico. In the early decades of the From Génico the path splits apart nineteenth century, a new military in two parts: following probably the medieval path, you can go to The most important feature of this Lierna. Instead you can head path is the equilibrium between inland, passing by the ancient human activities and the nature. church of Saint Peter in Ortanella. The Wayfarer’s trail is 40 Km long, Both routes meet at the castle of with difference in altitudes between Vezio, one of the most important 400 and 1000 meters. You can fortified hamlets of Eastern Lario. experience it in four stages. From Vezio the paths continue to The wayfarer’s trail is Bellano, where, after passing the characterized by orange signs Pioverna stream and its famous along the path and offers you the Horrid, you arrive at the sanctuary most attractive and evocative of Lezzeno. -

TOURIST GUIDE ASSOCIATIONS PROVINCE of COMO Associazione Guide E Accompagnatori Turistici Di Como E Provincia Phone No
TOURIST GUIDE www.lakecomo.com ISOLA COMACINA 01_ING_presen_sistema.indd 1 25/07/11 11:40 PRESENTATION This Tourist Guide introduces one of the most beautiful areas in the region called Lombardy and enthusiastically welcomes all visitors who are planning to have an enjoyable stay here. Seen from above, the blue of the lakes and the green of the woods are the two colours which exist in harmony in this spectacular landscape full of panoramas. The lakes are the main characteristic of Como and Lecco provinces, surrounded by a range of important mountains which open up to the hilly countryside of Brianza to the South, the home to entrepreneurship. We had the idea of preparing a guide that was not only easy to use, but of high quality: therefore, you will fi nd, alongside the usual cultural itineraries that inform you of our national heritage, practical information that can help you to easily discover our region and even the less known Via Sirtori 5 - 22100 Como places. Phone No. + 39 031 2755551 Subdivided into geographical areas of lake, mountain Fax + 39 031 2755569 and plain, the Guide describes the entire territory of [email protected] www.provincia.como.it Como and Lecco provinces; its history, architecture, art www.lakecomo.com and natural beauty, starting from the “capoluoghi” (main towns) of the province and the lake basin. It then goes on describing the mountain area and cultural features, uncovering the towns and ancient villages, alongside the mountain shelters and peaks. It gives detailed information on walking excursions for all nature lovers, from trekking to all types of sport. -

Sentiero Del Viandante
Attilio Premoli Il Sentiero del Viandante Quattro tappe tra Abbadia Lariana e Colico ripercorrendo le strade agricole ed i sentieri della sponda lecchese del lago Attilio Premoli L'ITINERARIO E LE TAPPE Il Sentiero del Viandante, sistemato e segnalato dall’Azienda di Promozione Turistica del Lecchese nel 1992, percorre la sponda orientale del lago di Como e unisce Abbadia Lariana al Santuario della Madonna di Valpozzo, nel comune di Piantedo, alle porte della Valtellina. Non parte da Lecco perché i lavori di costruzione della ferrovia e della strada provinciale hanno spazzato via ogni traccia dell’antico percorso. L’itinerario ha una lunghezza di circa 45 km e si può percorrere in tre o quattro tappe (ma anche, forzando la marcia, in due) per un totale di 16/17 ore. La suddivisione in quattro tappe, è certamente quella meno sportiva, ma consente di dedicare del tempo anche ad altro, non solo al puro esercizio del camminare. Lungo il percorso si incontrano infatti interessanti testimonianze storiche (chiese, fortificazioni, opere d’arte, ville, giardini, vecchi borghi ben conservati), ma anche fenomeni naturali (il Fiumelatte, l’Orrido di Bellano) assolutamente meritevoli di una visita non superficiale. Il Sentiero è suddiviso in quattro tappe, abbastanza omogenee sia sui chilometri da percorrere che sui dislivelli; la più difficoltosa può essere considerata la seconda tappa, quella che da Lierna ci porta fino a Varenna. Il dislivello se si privilegia la variante alta è di circa 990 mt. e lo si tocca presso la chiesetta di san Pietro uno dei luoghi più panoramici di tutto il Sentiero del Viandante e punto culminante della salita. -

App Differenziati E’ Facile, Intuitivo E Immediato Fornire Le Seguenti Informazioni E Servizi
Valmadrera, 27 novembre 2015 DA OGGI I CITTADINI DEI COMUNI DELLA PROVINCIA DI LECCO CHE HANNO AFFIDATO A SILEA IL SERVIZIO DI RACCOLTA DEI RIFIUTI, AVRANNO SEMPRE A DISPOSIZIONE UNA GUIDA MOBILE SEMPLICE DA CONSULTARE E AGGIORNATA, CON ACCESSO IMMEDIATO ALLE INFORMAZIONI SUI SERVIZI LEGATI AL MONDO DELL’IGIENE URBANA. Servizio di raccolta già avviato Servizio in avvio nel 2016 COMUNI E NUMERO DI ABITANTI INTERESSATI DAL SERVIZIO 1 ABBADIA LARIANA 3.227 38 LOMAGNA 4.952 2 ANNONE DI BRIANZA 2.346 39 MANDELLO DEL LARIO 10.421 3 BALLABIO 4.083 40 MERATE 14.868 4 BARZAGO 2.471 41 MISSAGLIA 8.751 5 BARZANO' 5.143 42 MOGGIO 488 6 BARZIO 1.301 43 MONTE MARENZO 1.958 7 BELLANO 3.248 44 MONTEVECCHIA 2.607 8 BOSISIO PARINI 3.476 45 MORTERONE 38 9 CALCO 5.255 46 NIBIONNO 3.704 10 CALOLZIOCORTE 14.112 47 OGGIONO 8.928 11 CARENNO 1.482 48 OLGIATE MOLGORA 6.376 12 CASATENOVO 12.934 49 OLGINATE 7.119 13 CASSINA VALSASSINA 482 50 OLIVETO LARIO 1.195 14 CASTELLO DI BRIANZA 2.557 51 PADERNO D'ADDA 3.927 15 CERNUSCO LOMBARDONE 3.825 52 PAGNONA 386 16 CESANA BRIANZA 2.410 53 PARLASCO 139 17 CIVATE 3.929 54 PASTURO 2.014 18 COLICO 7.719 55 PERLEDO 970 19 CORTENOVA 1.214 56 PESCATE 2.149 20 COSTA MASNAGA 4.801 57 PREMANA 2.293 21 CREMELLA 1.770 58 PRIMALUNA 2.244 22 CREMENO 1.482 59 PUSIANO 1.344 23 DERVIO 2.679 60 ROBBIATE 6.256 24 DOLZAGO 2.428 25 DORIO 314 61 ROGENO 3.194 26 ELLO 1.230 62 SANTA MARIA HOE' 2.205 27 ERVE 726 63 SIRONE 2.404 28 ESINO LARIO 764 64 SIRTORI 2.870 29 GALBIATE 8.581 65 SUEGLIO 144 30 GARBAGNATE MONASTERO 2.448 66 SUELLO 1.725 31