Light Induced Changes in Starry Flounder (Platichthys Stellatus) Opsin Expression 2 and Its Influence on Vision Estimated from a Camouflage-Based Behavioural Assay
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Practical Handbook for Determining the Ages of Gulf of Mexico And
A Practical Handbook for Determining the Ages of Gulf of Mexico and Atlantic Coast Fishes THIRD EDITION GSMFC No. 300 NOVEMBER 2020 i Gulf States Marine Fisheries Commission Commissioners and Proxies ALABAMA Senator R.L. “Bret” Allain, II Chris Blankenship, Commissioner State Senator District 21 Alabama Department of Conservation Franklin, Louisiana and Natural Resources John Roussel Montgomery, Alabama Zachary, Louisiana Representative Chris Pringle Mobile, Alabama MISSISSIPPI Chris Nelson Joe Spraggins, Executive Director Bon Secour Fisheries, Inc. Mississippi Department of Marine Bon Secour, Alabama Resources Biloxi, Mississippi FLORIDA Read Hendon Eric Sutton, Executive Director USM/Gulf Coast Research Laboratory Florida Fish and Wildlife Ocean Springs, Mississippi Conservation Commission Tallahassee, Florida TEXAS Representative Jay Trumbull Carter Smith, Executive Director Tallahassee, Florida Texas Parks and Wildlife Department Austin, Texas LOUISIANA Doug Boyd Jack Montoucet, Secretary Boerne, Texas Louisiana Department of Wildlife and Fisheries Baton Rouge, Louisiana GSMFC Staff ASMFC Staff Mr. David M. Donaldson Mr. Bob Beal Executive Director Executive Director Mr. Steven J. VanderKooy Mr. Jeffrey Kipp IJF Program Coordinator Stock Assessment Scientist Ms. Debora McIntyre Dr. Kristen Anstead IJF Staff Assistant Fisheries Scientist ii A Practical Handbook for Determining the Ages of Gulf of Mexico and Atlantic Coast Fishes Third Edition Edited by Steve VanderKooy Jessica Carroll Scott Elzey Jessica Gilmore Jeffrey Kipp Gulf States Marine Fisheries Commission 2404 Government St Ocean Springs, MS 39564 and Atlantic States Marine Fisheries Commission 1050 N. Highland Street Suite 200 A-N Arlington, VA 22201 Publication Number 300 November 2020 A publication of the Gulf States Marine Fisheries Commission pursuant to National Oceanic and Atmospheric Administration Award Number NA15NMF4070076 and NA15NMF4720399. -

The Osmoregulatory Metabolism Op the Starry Flounder, Platichthys Stellatus
THE OSMOREGULATORY METABOLISM OP THE STARRY FLOUNDER, PLATICHTHYS STELLATUS by CLEVELAND PENDLETON HICKMAN, JR. B.A., DePauw University, 1950 M.S., University of New Hampshire, 1953 A THESIS SUBMITTED IN PARTIAL FULFILMENT OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY in the Department of Zoology We accept this thesis as conforming to the required standard. THE UNIVERSITY OF BRITISH COLUMBIA June, 1958 Faculty of Graduate Studies PROGRAMME OF THE FINAL ORAL EXAMINATION FOR THE DEGREE OF DOCTOR OF PHILOSOPHY of CLEVELAND PENDLETON HICKMAN JR. B.A. DePauw University, 1950 M.S. University of New Hampshire, 1953 IN ROOM 187A, BIOLOGICAL SCIENCES BUILDING MONDAY, JUNE 30, 1958 at 10:30 a.m. COMMITTEE IN CHARGE DEAN F. H. SOWARD, Chairman H. ADASKIN W. S. HOAR W. A. CLEMENS W. N. HOLMES I. McT. COWAN C. C. LINDSEY P. A. DEHNEL H. McLENNAN R. F. SCAGEL External Examiner: F. E. J. FRY University of Toronto THE OSMOREGULATORY METABOLISM OF THE STARRY FLOUNDER, PLATICHTYS STELLATUS ABSTRACT Energy demands for osmotic regulation and the possible osmoregulatory role of the thyroid gland were investigated in the euryhaline starry flounder, Platichthys stellatus. Using a melt• ing-point technique, it was established that flounder could regulate body fluid concentration independent of widely divergent environ• mental salinities. Small flounder experienced more rapid disturb• ances of body fluid concentration than large flounder after abrupt salinity alterations. The standard metabolic rate of flounder adapted to fresh water was consistently and significantly less than that of marine flounder. In supernormal salinities standard metabolic rate was significantly greater than in normal sea water. -

Preliminary Mass-Balance Food Web Model of the Eastern Chukchi Sea
NOAA Technical Memorandum NMFS-AFSC-262 Preliminary Mass-balance Food Web Model of the Eastern Chukchi Sea by G. A. Whitehouse U.S. DEPARTMENT OF COMMERCE National Oceanic and Atmospheric Administration National Marine Fisheries Service Alaska Fisheries Science Center December 2013 NOAA Technical Memorandum NMFS The National Marine Fisheries Service's Alaska Fisheries Science Center uses the NOAA Technical Memorandum series to issue informal scientific and technical publications when complete formal review and editorial processing are not appropriate or feasible. Documents within this series reflect sound professional work and may be referenced in the formal scientific and technical literature. The NMFS-AFSC Technical Memorandum series of the Alaska Fisheries Science Center continues the NMFS-F/NWC series established in 1970 by the Northwest Fisheries Center. The NMFS-NWFSC series is currently used by the Northwest Fisheries Science Center. This document should be cited as follows: Whitehouse, G. A. 2013. A preliminary mass-balance food web model of the eastern Chukchi Sea. U.S. Dep. Commer., NOAA Tech. Memo. NMFS-AFSC-262, 162 p. Reference in this document to trade names does not imply endorsement by the National Marine Fisheries Service, NOAA. NOAA Technical Memorandum NMFS-AFSC-262 Preliminary Mass-balance Food Web Model of the Eastern Chukchi Sea by G. A. Whitehouse1,2 1Alaska Fisheries Science Center 7600 Sand Point Way N.E. Seattle WA 98115 2Joint Institute for the Study of the Atmosphere and Ocean University of Washington Box 354925 Seattle WA 98195 www.afsc.noaa.gov U.S. DEPARTMENT OF COMMERCE Penny. S. Pritzker, Secretary National Oceanic and Atmospheric Administration Kathryn D. -

Humboldt Bay Fishes
Humboldt Bay Fishes ><((((º>`·._ .·´¯`·. _ .·´¯`·. ><((((º> ·´¯`·._.·´¯`·.. ><((((º>`·._ .·´¯`·. _ .·´¯`·. ><((((º> Acknowledgements The Humboldt Bay Harbor District would like to offer our sincere thanks and appreciation to the authors and photographers who have allowed us to use their work in this report. Photography and Illustrations We would like to thank the photographers and illustrators who have so graciously donated the use of their images for this publication. Andrey Dolgor Dan Gotshall Polar Research Institute of Marine Sea Challengers, Inc. Fisheries And Oceanography [email protected] [email protected] Michael Lanboeuf Milton Love [email protected] Marine Science Institute [email protected] Stephen Metherell Jacques Moreau [email protected] [email protected] Bernd Ueberschaer Clinton Bauder [email protected] [email protected] Fish descriptions contained in this report are from: Froese, R. and Pauly, D. Editors. 2003 FishBase. Worldwide Web electronic publication. http://www.fishbase.org/ 13 August 2003 Photographer Fish Photographer Bauder, Clinton wolf-eel Gotshall, Daniel W scalyhead sculpin Bauder, Clinton blackeye goby Gotshall, Daniel W speckled sanddab Bauder, Clinton spotted cusk-eel Gotshall, Daniel W. bocaccio Bauder, Clinton tube-snout Gotshall, Daniel W. brown rockfish Gotshall, Daniel W. yellowtail rockfish Flescher, Don american shad Gotshall, Daniel W. dover sole Flescher, Don stripped bass Gotshall, Daniel W. pacific sanddab Gotshall, Daniel W. kelp greenling Garcia-Franco, Mauricio louvar -

UC San Diego Previously Published Works
UC San Diego UC San Diego Previously Published Works Title Differential effects of bicarbonate on severe hypoxia- and hypercapnia-induced cardiac malfunctions in diverse fish species. Permalink https://escholarship.org/uc/item/8p99n97z Journal Journal of comparative physiology. B, Biochemical, systemic, and environmental physiology, 191(1) ISSN 0174-1578 Authors Lo, Mandy Shahriari, Arash Roa, Jinae N et al. Publication Date 2021 DOI 10.1007/s00360-020-01324-y Peer reviewed eScholarship.org Powered by the California Digital Library University of California 1 Differential effects of bicarbonate on severe hypoxia- and hypercapnia-induced cardiac 2 malfunctions in diverse fish species 3 4 Mandy Lo1, Arash Shahriari1, Jinae N. Roa2, Martin Tresguerres2, Anthony P. Farrell1,3 5 6 7 1Department of Zoology, University of British Columbia, 6270 University Boulevard, 8 Vancouver, British Columbia, Canada V6T 1Z4 9 10 2Marine Biology Research Division, Scripps Institution of Oceanography, University of 11 California San Diego, 9500 Gilman Drive, La Jolla, CA 92093, USA 12 13 3Faculty of Land and Food Systems, University of British Columbia, 2357 Main Mall, 14 Vancouver, British Columbia, Canada V6T 1Z4 15 16 M. Lo (corresponding author) 17 Department of Zoology, University of British Columbia, 6270 University Boulevard, Vancouver, 18 British Columbia, Canada V6T 1Z4 19 Email: [email protected] 20 21 22 23 1 24 Abstract 25 We tested in six fish species [Pacific lamprey (Lampetra richardsoni), Pacific spiny dogfish 26 (Squalus suckleyi), Asian swamp eel (Monopterus albus), white sturgeon (Acipenser 27 transmontanus), zebrafish (Danio rerio), and starry flounder (Platichthys stellatus)] the - 28 hypothesis that elevated extracellular [HCO3 ] protects spontaneous heart rate and cardiac force 29 development from the known impairments that severe hypoxia and hypercapnic acidosis can 30 induce. -

Flounders, Halibuts, Soles Capture Production by Species, Fishing Areas
101 Flounders, halibuts, soles Capture production by species, fishing areas and countries or areas B-31 Flets, flétans, soles Captures par espèces, zones de pêche et pays ou zones Platijas, halibuts, lenguados Capturas por especies, áreas de pesca y países o áreas Species, Fishing area Espèce, Zone de pêche 2009 2010 2011 2012 2013 2014 2015 2016 2017 2018 Especie, Área de pesca t t t t t t t t t t Mediterranean scaldfish Arnoglosse de Méditerranée Serrandell Arnoglossus laterna 1,83(01)001,01 MSF 34 Italy - - - - - - - 57 223 123 34 Fishing area total - - - - - - - 57 223 123 37 Italy ... ... ... ... ... ... 447 479 169 403 37 Fishing area total ... ... ... ... ... ... 447 479 169 403 Species total ... ... ... ... ... ... 447 536 392 526 Leopard flounder Rombou léopard Lenguado leopardo Bothus pantherinus 1,83(01)018,05 OUN 51 Bahrain 2 - - 1 1 4 4 F 4 F 4 F 4 F Saudi Arabia 77 80 77 75 74 83 71 79 80 F 74 51 Fishing area total 79 80 77 76 75 87 75 F 83 F 84 F 78 F Species total 79 80 77 76 75 87 75 F 83 F 84 F 78 F Lefteye flounders nei Arnoglosses, rombous nca Rodaballos, rombos, etc. nep Bothidae 1,83(01)XXX,XX LEF 21 USA 1 087 774 566 747 992 759 545 406 633 409 21 Fishing area total 1 087 774 566 747 992 759 545 406 633 409 27 Germany - - - - - - - - 0 - Portugal 136 103 143 125 105 102 87 76 84 105 Spain 134 116 96 56 29 8 12 12 6 5 27 Fishing area total 270 219 239 181 134 110 99 88 90 110 31 USA 59 38 71 45 41 128 117 133 99 102 31 Fishing area total 59 38 71 45 41 128 117 133 99 102 34 Greece - - - - - - - 71 45 - Portugal 15 46 .. -

Seamap Environmental and Biological Atlas of the Gulf of Mexico, 2014
environmental and biological atlas of the gulf of mexico 2014 gulf states marine fisheries commission number 262 february 2017 seamap SEAMAP ENVIRONMENTAL AND BIOLOGICAL ATLAS OF THE GULF OF MEXICO, 2014 Edited by Jeffrey K. Rester Gulf States Marine Fisheries Commission Manuscript Design and Layout Ashley P. Lott Gulf States Marine Fisheries Commission GULF STATES MARINE FISHERIES COMMISSION FEBRUARY 2017 NUMBER 262 This project was supported in part by the National Oceanic and Atmospheric Administration, National Marine Fisheries Service, under State/Federal Project Number NA16NMFS4350111. GULF STATES MARINE FISHERIES COMMISSION COMMISSIONERS ALABAMA John Roussel N. Gunter Guy, Jr. 1221 Plains Port Hudson Road Alabama Department of Conservation Zachary, LA 70791 and Natural Resources 64 North Union Street MISSISSIPPI Montgomery, AL 36130-1901 Jamie Miller, Executive Director Mississippi Department of Marine Resources Steve McMillan 1141 Bayview Avenue P.O. Box 337 Biloxi, MS 39530 Bay Minette, AL 36507 Senator Brice Wiggins Chris Nelson 1501 Roswell Street Bon Secour Fisheries, Inc. Pascagoula, MS 39581 P.O. Box 60 Bon Secour, AL 36511 Joe Gill, Jr. Joe Gill Consulting, LLC FLORIDA 910 Desoto Street Nick Wiley, Executive Director Ocean Springs, MS 39566-0535 FL Fish and Wildlife Conservation Commission 620 South Meridian Street TEXAS Tallahassee, FL 32399-1600 Carter Smith, Executive Director Texas Parks and Wildlife Department Senator Thad Altman 4200 Smith School Road State Senator, District 24 Austin, TX 78744 6767 North Wickham Road, Suite 211 Melbourne, FL 32940 Troy B. Williamson, II P.O. Box 967 TBA Corpus Christi, TX 78403 LOUISIANA Representative Wayne Faircloth Jack Montoucet, Secretary Texas House of Representatives LA Department of Wildlife and Fisheries 2121 Market Street, Suite 205 P.O. -

8. Biological Resources March 5, 2003 Page 8-1
Marina Shores Village Project Draft EIR City of Redwood City 8. Biological Resources March 5, 2003 Page 8-1 8. BIOLOGICAL RESOURCES This EIR chapter describes the existing biological resources on the Marina Shores Village project site and in the project vicinity, potential project impacts on those resources, and identifies measures warranted to mitigate identified significant impacts. The descriptions in this chapter are based on an independent evaluation conducted by the EIR consulting biologist, Wetlands Research Associates, based on field reconnaissance of the project site; review of a biological assessment and preliminary wetlands investigation and delineation prepared for the applicant by the Huffman-Broadway Group, Inc.;1 review of the most current versions of the California Natural Diversity Data Base (2002); and review of pertinent biological resources publications and data available from other sources, including the U.S. Fish and Wildlife Service (USFWS), U.S. Environmental Protection Agency (USEPA), National Marine Fisheries Service (NMFS), U.S. Army Corps of Engineers (Corps), California Department of Fish and Game (CDFG), San Francisco Bay Regional Water Quality Control Board (RWQCB), Sequoia Audubon Society, and California Exotic Pest Plan Council (see section 8.4, References, herein). 8.1 SETTING The project site is located in an area that has undergone significant human disturbance, including filling, vegetation removal, soil compaction, and altered runoff patterns.2 Five habitat 1The Huffman-Broadway Group, Inc., 2002a. Biological Assessment: Marina Shores Village Project, Redwood City, California. Larkspur, California. Prepared for Glenborough-Pauls, LLC, February 2002. Also, The Huffman-Broadway Group, Inc., 2002b. Investigation of the Presence of Wetlands and Other Waters of the United States on the Marina Shores Village Project Site, Redwood City, California. -

Sand Flounder (Family Paralichthyidae) Diversity in North Carolina by the Ncfishes.Com Team
Sand Flounder (Family Paralichthyidae) Diversity in North Carolina By the NCFishes.com Team Along North Carolina’s shore there are four families of flatfish comprising 36 species having eyes on the left side of their body facing upward when lying in or atop the substrate (NCFishes.com; Table 1; Figure 1). The families and species can be confusing to tell apart. The key characteristics provided for in Table 1 should enable one to differentiate between the four families and this document will aid you in the identification of the species in the Family Paralichthyidae in North Carolina. Table 1. The four families of left-facing flounders found along and off the coast of North Carolina. Family Common Name Key Characteristics (adapted from Munroe 2002) No. Species Preopercle exposed, its posterior margin free and visible, not hidden by skin or scales. Dorsal fin long, originating above, lateral to, or anterior to upper eye. Dorsal and anal fins not attached to caudal fin. Both pectoral Paralichthyidae Sand Flounders fins present. Both pelvic fins present, with 5 or 6 rays. 20 Margin of preopercle not free (hidden beneath skin and scales). Pectoral fins absent in adults. Lateral line absent on both sides of body. Cynoglossidae Tonguefishes Dorsal and anal fins joined to caudal fin. No branched caudal-fin rays. 9 Lateral line absent or poorly developed on blind side; lateral line absent below lower eye. Lateral line of eyed side with high arch over pectoral Bothidae Lefteye Flounders fin. Pelvic fin of eyed side on midventral line. 6 Both pelvic fins elongate, placed close to midline and extending forward to urohyal. -

The Flounder Fishery of the Gulf of Mexico, United States: a Regional Management Plan
The Flounder Fishery of the Gulf of Mexico, United States: A Regional Management Plan ..... .. ·. Gulf States Marine Fisheries Commission October 2000 Number83 GULF STATES MARINE FISHERIES COMMISSION Commissioners and Proxies Alabama Warren Triche Riley Boykin Smith Louisiana House of Representatives Alabama Department of Conservation & Natural 100 Tauzin Lane Resources Thibodaux, Louisiana 70301 64 North Union Street Montgomery, Alabama 36130-1901 Frederic L. Miller proxy: Vernon Minton P.O. Box 5098 Marine Resources Division Shreveport, Louisiana 71135-5098 P.O. Drawer 458 Gulf Shores, Alabama 36547 Mississippi Glenn H. Carpenter Walter Penry Mississippi Department of Marine Resources Alabama House of Representatives 1141 Bayview Avenue, Suite 101 12040 County Road 54 Biloxi, Mississippi 39530 Daphne, Alabama 36526 proxy: William S. “Corky” Perret Mississippi Department of Marine Resources Chris Nelson 1141 Bayview Avenue, Suite 101 Bon Secour Fisheries, Inc. Biloxi, Mississippi 39530 P.O. Box 60 Bon Secour, Alabama 36511 Billy Hewes Mississippi Senate Florida P.O. Box 2387 Allan L. Egbert Gulfport, Mississippi 39505 Florida Fish & Wildlife Conservation Commission 620 Meridian Street George Sekul Tallahassee, Florida 323299-1600 805 Beach Boulevard, #302 proxies: Ken Haddad, Director Biloxi, Mississippi 39530 Florida Marine Research Institute 100 Eighth Avenue SE Texas St. Petersburg, Florida 33701 Andrew Sansom Texas Parks & Wildlife Department Ms. Virginia Vail 4200 Smith School Road Division of Marine Resources Austin, Texas 78744 Fish & Wildlife Conservation Commission proxies: Hal Osburn and Mike Ray 620 Meridian Street Texas Parks & Wildlife Department Tallahassee, Florida 32399-1600 4200 Smith School Road Austin, Texas 78744 William W. Ward 2221 Corrine Street J.E. “Buster” Brown Tampa, Florida 33605 Texas Senate P.O. -
St. Lucie, Units 1 and 2
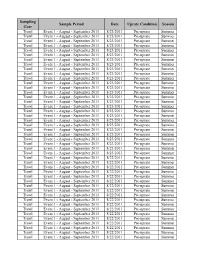
Sampling Sample Period Date Uprate Condition Season Gear Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event -

Kosher Fish List.Xlsx
Below is a comprehensive list of Kosher fish. A Albacore See: Mackerel Alewife See: Herring Amberjack See: Jack Anchovies (Family Engraulidae). Including: European anchovy (Engraulis encrasciolus), North Angelfish and butterfly fish (Family Chaetodontidae). Including: Angelfish (Holacanthus Atlantic Pomfret or Ray's Bream (Brama brama) B Ballyhoo See: Flyingfish Barracuda (Family Sphyraenidae) Including: Barracuda and kakus (Sphyraena species). Bass, Sea Bass Temperate bass, Sunfish, Drums Bigeye (Family Priacanthidae). Including: Bigeyes or aweoweos (Priacanthus species). Blackfish See: Carp, Wrass Blacksmith See: Damselfish Blueback See: Flounder, Herring, Trout Bluefish or snapper blue (Pomatomus saltarix) Bluegill See: Sunfish Bocaccio See: Scorpionfish Bombay duck (Harpadeon nehereus) Bonefish (Albula vulpes) Bonito See: Cobia, Mackerel Bowfin Freshwater dogfish, or grindle (Amia calva) Bream See: Carp, Atlantic pomfret, Porgie Brill See: Flounder Buffalo fish See: Sucker ButterFish (Family Stromateidae), Including: Butterfish (Peprilus tracanthus); Pacific pompano (Peprilus similimus); harvestfish (Peprilus species) Butterfly fish See: Angelfish C http://www.kashrut.com/articles/fish/ Cabrilla See: Sea Bass Calico bass See: Sunfish Capelin See: Smelt Carp and minnow (Family Cyprinidae), Including: the carp, leather carp, mirror carp (Cyprinus carpio); Crucian carp (Carassius carassius); Goldfish (Carassius auratus); tench (Tinca tinca); Splittail (Pogonichthys macrolepidotus); Squawfish (Ptychocheilus species); Scramento Carosucker