Two New Cavernicolous Genera of Julidae (Diplopoda, Julida), with Notes on the Tribe Brachyiulini and on Julid Subanal Hooks and Anchors
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Cylindroiulus Truncorum (Silvestri): a New Milliped for Virginia (USA), with Natural History Observations (Julida: Julidae)
Banisteria, Number 20, 2002 © 2002 by the Virginia Natural History Society Cylindroiulus truncorum (Silvestri): A New Milliped for Virginia (USA), with Natural History Observations (Julida: Julidae) Jorge A. Santiago-Blay Department of Paleobiology, MRC-121 National Museum of Natural History 10th and Constitution Avenue Smithsonian Institution P.O. Box 37012 Washington, DC 20013-7012 Richard L. Hoffman Virginia Museum of Natural History Martinsville, Virginia 24112 Joseph B. Lambert and Yuyang Wu Department of Chemistry Northwestern University 2145 Sheridan Road Evanston, Illinois 60208-3113 INTRODUCTION truncorum for Raleigh, North Carolina, about 320 km SSE of Salem (Shelley, 1978) is the southernmost In the fall 2000, author SB cleared the underbrush known occurrence of this species in the United States. of an Eastern White Pine (Pinus strobus L.) grove in his This milliped has also been documented for Brazil backyard located in an urban area of Salem, Virginia (Chamberlin & Hoffman, 1958; Hoffman, 1999). (USA) by cutting and removing the lower branches. About a year later, he revisited the same trees and Natural History Observations noticed copious resinous exudations originating from the branch stumps, particularly on five of the trees. Berlese extractions from P. strobus leaf litter were There, he observed about twenty millipeds, later conducted in November 2001 and yielded a maximum identified as Cylindroiulus truncorum (Silvestri, 1896; of about 50 C. truncorum per 0.25 m2 (= 200 C. species group reviewed by Korsós & Enghoff, 1990), truncorum per m2). In his many years of studying soil attached to the resin, 1-2 meters above ground (Fig. 1). invertebrates and running numerous Berlese samples, Voucher specimens of Cylindroiulus truncorum are particularly in southwestern Virginia, RLH has seldom deposited at the Virginia Museum of Natural History encountered millipeds under pine litter. -
Subterranean Biodiversity and Depth Distribution of Myriapods in Forested Scree Slopes of Central Europe
A peer-reviewed open-access journal ZooKeys Subterranean930: 117–137 (2020) biodiversity and depth distribution of myriapods in forested scree slopes of... 117 doi: 10.3897/zookeys.930.48914 RESEARCH ARTICLE http://zookeys.pensoft.net Launched to accelerate biodiversity research Subterranean biodiversity and depth distribution of myriapods in forested scree slopes of Central Europe Beáta Haľková1, Ivan Hadrián Tuf 2, Karel Tajovský3, Andrej Mock1 1 Institute of Biology and Ecology, Faculty of Science, Pavol Jozef Šafárik University, Košice, Slovakia 2 De- partment of Ecology and Environmental Sciences, Faculty of Science, Palacky University, Olomouc, Czech Republic 3 Institute of Soil Biology, Biology Centre CAS, České Budějovice, Czech Republic Corresponding author: Beáta Haľková ([email protected]) Academic editor: L. Dányi | Received 28 November 2019 | Accepted 10 February 2020 | Published 28 April 2020 http://zoobank.org/84BEFD1B-D8FA-4B05-8481-C0735ADF2A3C Citation: Haľková B, Tuf IH, Tajovský K, Mock A (2020) Subterranean biodiversity and depth distribution of myriapods in forested scree slopes of Central Europe. In: Korsós Z, Dányi L (Eds) Proceedings of the 18th International Congress of Myriapodology, Budapest, Hungary. ZooKeys 930: 117–137. https://doi.org/10.3897/zookeys.930.48914 The paper is dedicated to Christian Juberthie (12 Mar 1931–7 Nov 2019), the author of the concept of MSS (milieu souterrain superficiel) and the doyen of modern biospeleology Abstract The shallow underground of rock debris is a unique animal refuge. Nevertheless, the research of this habitat lags far behind the study of caves and soil, due to technical and time-consuming demands. Data on Myriapoda in scree habitat from eleven localities in seven different geomorphological units of the Czech and Slovak Republics were processed. -

Millipedes (Diplopoda) from Caves of Portugal
A.S.P.S. Reboleira and H. Enghoff – Millipedes (Diplopoda) from caves of Portugal. Journal of Cave and Karst Studies, v. 76, no. 1, p. 20–25. DOI: 10.4311/2013LSC0113 MILLIPEDES (DIPLOPODA) FROM CAVES OF PORTUGAL ANA SOFIA P.S. REBOLEIRA1 AND HENRIK ENGHOFF2 Abstract: Millipedes play an important role in the decomposition of organic matter in the subterranean environment. Despite the existence of several cave-adapted species of millipedes in adjacent geographic areas, their study has been largely ignored in Portugal. Over the last decade, intense fieldwork in caves of the mainland and the island of Madeira has provided new data about the distribution and diversity of millipedes. A review of millipedes from caves of Portugal is presented, listing fourteen species belonging to eight families, among which six species are considered troglobionts. The distribution of millipedes in caves of Portugal is discussed and compared with the troglobiont biodiversity in the overall Iberian Peninsula and the Macaronesian archipelagos. INTRODUCTION All specimens from mainland Portugal were collected by A.S.P.S. Reboleira, while collectors of Madeiran speci- Millipedes play an important role in the decomposition mens are identified in the text. Material is deposited in the of organic matter, and several species around the world following collections: Zoological Museum of University of have adapted to subterranean life, being found from cave Copenhagen, Department of Animal Biology, University of entrances to almost 2000 meters depth (Culver and Shear, La Laguna, Spain and in the collection of Sofia Reboleira, 2012; Golovatch and Kime, 2009; Sendra and Reboleira, Portugal. 2012). Although the millipede faunas of many European Species were classified according to their degree of countries are relatively well studied, this is not true of dependence on the subterranean environment, following Portugal. -

Studies of the Laboulbeniomycetes: Diversity, Evolution, and Patterns of Speciation
Studies of the Laboulbeniomycetes: Diversity, Evolution, and Patterns of Speciation The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters Citable link http://nrs.harvard.edu/urn-3:HUL.InstRepos:40049989 Terms of Use This article was downloaded from Harvard University’s DASH repository, and is made available under the terms and conditions applicable to Other Posted Material, as set forth at http:// nrs.harvard.edu/urn-3:HUL.InstRepos:dash.current.terms-of- use#LAA ! STUDIES OF THE LABOULBENIOMYCETES: DIVERSITY, EVOLUTION, AND PATTERNS OF SPECIATION A dissertation presented by DANNY HAELEWATERS to THE DEPARTMENT OF ORGANISMIC AND EVOLUTIONARY BIOLOGY in partial fulfillment of the requirements for the degree of Doctor of Philosophy in the subject of Biology HARVARD UNIVERSITY Cambridge, Massachusetts April 2018 ! ! © 2018 – Danny Haelewaters All rights reserved. ! ! Dissertation Advisor: Professor Donald H. Pfister Danny Haelewaters STUDIES OF THE LABOULBENIOMYCETES: DIVERSITY, EVOLUTION, AND PATTERNS OF SPECIATION ABSTRACT CHAPTER 1: Laboulbeniales is one of the most morphologically and ecologically distinct orders of Ascomycota. These microscopic fungi are characterized by an ectoparasitic lifestyle on arthropods, determinate growth, lack of asexual state, high species richness and intractability to culture. DNA extraction and PCR amplification have proven difficult for multiple reasons. DNA isolation techniques and commercially available kits are tested enabling efficient and rapid genetic analysis of Laboulbeniales fungi. Success rates for the different techniques on different taxa are presented and discussed in the light of difficulties with micromanipulation, preservation techniques and negative results. CHAPTER 2: The class Laboulbeniomycetes comprises biotrophic parasites associated with arthropods and fungi. -

Diplopoda, Julidae) in the Northern Atlanticinternat Region...Ional Journal69 of Doi: 10.3897/Ijm.7.3064 Data Paper Myriapodology
IJM 7: 62–91 (2012) A peer-reviewed open-access journal Phoretic mite associates of millipedes (Diplopoda, Julidae) in the northern AtlanticINTERNAT region...IONAL JOURNAL69 OF doi: 10.3897/ijm.7.3064 DATA PAPER www.pensoft.net/journals/ijm Myriapodology Phoretic mite associates of millipedes (Diplopoda, Julidae) in the northern Atlantic region (North America, Europe) Monica A. Farfan1,2, Hans Klompen2 1 Acarology Laboratory, Ohio State University, Columbus, Ohio, 43212, USA 2 Dept. of Biological Sciences, Univ. Illinois at Chicago, Chicago, Illinois, 60607, USA Corresponding author: Monica Farfan ([email protected]) Academic editor: Pavel Stoev | Received 10 March 2012 | Accepted 22 May 2012 | Published 5 June 2012 Citation: Farfan MA, Klompen H (2012) Phoretic mite associates of millipedes (Diplopoda, Julidae) in the northern Atlantic region (North America, Europe). International Journal of Myriapodology 7: 69–91. doi: 10.3897/ijm.7.3064 Abstract Introduced millipede species in the family Julidae are common in the U.S. but little is known about how they interact with other organisms, such as mites. To start to determine the nature of the relationship, millipedes were sampled from across the eastern U.S.A. and the United Kingdom in 2008–2009. Sixteen morphospecies of mites (Acari: Astigmata, Mesostigmata) were collected from these millipedes, 12 of which from a total of 13 species of julid millipedes. None of these 12 species was restricted to a single host species. However, 12 of the 16 mite species collected were restricted to either the U.S.A. or the U.K. These results are consistent with locality, rather than host, specificity. -

Gonopod Mechanics in Centrobolus Cook (Spirobolida: Trigoniulidae) II
Journal of Entomology and Zoology Studies 2016; 4(2): 152-154 E-ISSN: 2320-7078 P-ISSN: 2349-6800 JEZS 2016; 4(2): 152-154 Gonopod mechanics in Centrobolus Cook © 2016 JEZS (Spirobolida: Trigoniulidae) II. Images Received: 06-01-2016 Accepted: 08-02-2016 Mark Ian Cooper Mark Ian Cooper A) Department of Biological Sciences, Private Bag X3, Abstract University of Cape Town, Gonopod mechanics were described for four species of millipedes in the genus Centrobolus and are now Rondebosch 7701, South Africa. figured using scanning electron microscopy (SEM) with the aim to show the mechanism of sperm B) Electron Microscope Unit & Structural Biology Research competition. Structures of sperm displacement include projections on a moveable telopodite and tips on a Unit, University of Cape Town, distal process (opisthomerite). Three significant contact zones between the male and female genitalia South Africa. were recognized: (1) distal telopodite of the coleopod and the vulva, (2) phallopod and the bursa, (3) sternite and legs of the female. Keywords: coleopods, diplopod, gonopods, phallopods 1. Introduction The dual function of millipede male genitalia in sperm displacement and transfer were predicted from the combined examination of the ultrastructures of the male and female genitalia [1-3]. Genitalic structures function do not only in sperm transfer during the time of copulation, but that they perform copulatory courtship through movements and interactions with the female genitalia [4-5]. These 'functional luxuries' can induce cryptic female choice by stimulating structures on the female genitalia while facilitating rival-sperm displacement and sperm transfer. Genitalic complexity is probably underestimated in many species because they have only been studied in the retracted or relaxed state [4]. -

A New Genus of the Millipede Tribe Brachyiulini (Diplopoda: Julida: Julidae) from the Aegean Region
European Journal of Taxonomy 70: 1-12 ISSN 2118-9773 http://dx.doi.org/10.5852/ejt.2013.70 www.europeanjournaloftaxonomy.eu 2013 · Lazányi E. & Vagalinski B. This work is licensed under a Creative Commons Attribution 3.0 License. Research article urn:lsid:zoobank.org:pub:5E23F454-2A68-42F6-86FB-7D9952B2FE7B A new genus of the millipede tribe Brachyiulini (Diplopoda: Julida: Julidae) from the Aegean region Eszter LAZÁNYI1,4 & Boyan VAGALINSKI2,3,5 1 Corresponding author: Department of Zoology, Hungarian Natural History Museum, Baross u. 13, H-1088 Budapest, Hungary. E-mail: [email protected] 2 Faculty of Biology, Sofia University, 8 Dragan Tsankov Blvd., 1164 Sofia, Bulgaria. E-mail: [email protected] 3 Institute of Biodiversity and Ecosystem Research, Bulgarian Academy of Sciences, 2 Yurii Gagarin Street, 1113, Sofia, Bulgaria. 4 urn:lsid:zoobank.org:author:02DB48F1-624C-4435-AF85-FA87168CD85A 5 urn:lsid:zoobank.org:author:973B8725-039E-4F29-8D73-96A7F52CF934 Abstract. A new genus of the julid tribe Brachyiulini, Enghophyllum gen. nov., is described, comprising two species from Greece. The type-species, E. naxium (Verhoeff, 1901) comb. nov. (ex Megaphyllum Verhoeff, 1894), appears to be rather widespread in the Aegean: it is known from Antiparos Island and Naxos Island (the type locality), both in the Cyclades, as well as East Mavri Islet, Dodecanese Archipelago (new record). The vulva of E. naxium is described for the first time. In addition,E. sifnium gen. et sp. nov. is described based on a single adult male from Sifnos Island, Cyclades. The new genus is distinct from other genera of the Brachyiulini mainly by its peculiar gonopod structure, apparently disjunct and at least mostly apomorphous: (1) promeres broad, shield-like, in situ protruding mostly posteriad, completely covering the opisthomeres and gonopodal sinus; (2) transverse muscles and coxal apodemes of promere fully reduced; (3) opisthomere with three differentiated processes, i.e., lateral, basal posterior and apical posterior; (4) solenomere rather simple, tubular. -

Megaphyllum Unilineatum (C
RESEARCH ARTICLE Multifaceted activity of millipede secretions: Antioxidant, antineurodegenerative, and anti- Fusarium effects of the defensive secretions of Pachyiulus hungaricus (Karsch, 1881) and Megaphyllum unilineatum (C. L. Koch, 1838) (Diplopoda: Julida) a1111111111 1 2 2 2 a1111111111 Bojan IlićID *, Nikola Unković , Aleksandar KnezÏević , ZÏ eljko SavkovićID , Milica Ljaljević a1111111111 Grbić2, Jelena Vukojević2, Zvezdana Jovanović1, Slobodan Makarov1, Luka Lučić1 a1111111111 a1111111111 1 Department of Animal Development, University of BelgradeÐFaculty of Biology, Belgrade, Serbia, 2 Department of Algology, Mycology, and Lichenology, University of BelgradeÐFaculty of Biology, Belgrade, Serbia * [email protected] OPEN ACCESS Citation: Ilić B, Unković N, KnezÏević A, Savković ZÏ, Abstract Ljaljević Grbić M, Vukojević J, et al. (2019) Multifaceted activity of millipede secretions: Members of the millipede order Julida rely on dominantly quinonic defensive secretions with Antioxidant, antineurodegenerative, and anti- several minor, non-quinonic components. The free radical-scavenging activities of ethanol, Fusarium effects of the defensive secretions of methanol, hexane, and dichloromethane extracts of defensive secretions emitted by Pachyiulus hungaricus (Karsch, 1881) and Megaphyllum unilineatum (C. L. Koch, 1838) Pachyiulus hungaricus (Karsch, 1881) and Megaphyllum unilineatum (C. L. Koch, 1838) (Diplopoda: Julida). PLoS ONE 14(1): e0209999. were investigated using the ABTS, DPPH, and total reducing power (TRP) tests. The https://doi.org/10.1371/journal.pone.0209999 obtained extracts were also tested for inhibition of acetylcholinesterase and tyrosinase Editor: David A. Lightfoot, College of Agricultural activity. Finally, the antifungal potential of both julid extracts was evaluated against seven Sciences, UNITED STATES Fusarium species. Secretions of both species showed activity against free radicals, acetyl- Received: August 28, 2018 cholinesterase, tyrosinase, and all of the selected fungal species. -

Quinones and Non-Quinones from the Defensive Secretion of Unciger Transsilvanicus (Verhoeff, 1899) (Diplopoda, Julida, Julidae), from Serbia
Arch. Biol. Sci., Belgrade, 66 (1), 385-390, 2014 DOI:10.2298/ABS1401385S QUINONES AND NON-QUINONES FROM THE DEFENSIVE SECRETION OF UNCIGER TRANSSILVANICUS (VERHOEFF, 1899) (DIPLOPODA, JULIDA, JULIDAE), FROM SERBIA TATJANA LJ. SEKULIĆ1, LJ. V. VUJISIĆ2, B. P. M. ĆURČIĆ1, B. M. MANDIĆ2, D. Ž. ANTIĆ1, SNEŽANA S. TRIFUNOVIĆ2, D. M. GOĐEVAC3, VLATKA E. VAJS3, V. T. TOMIĆ1 and S. E. MAKAROV1 1 Institute of Zoology, Faculty of Biology, University of Belgrade, 11000 Belgrade, Serbia 2 Faculty of Chemistry, University of Belgrade, 11000 Belgrade, Serbia 3 Institute of Chemistry, Technology and Metallurgy, University of Belgrade, 11000 Belgrade, Serbia Abstract – A complex mixture of compounds was identified from the secretion of specimens of Unciger transsilvanicus. Phenol and p-cresol were detected for the first time in the family Julidae, and for the second time in the order Julida. Thirteen quinones were identified, with a great relative abundance of toloquinone and 2-methoxy-3-methyl-1,4-benzoqui- none. Hydroquinone was detected for the first time in the order Julida. Besides these compounds, isopentyl hexacosatetra- enoate and isopentyl esters of saturated and unsaturated fatty acids with chain lengths from C14 to C20were identified. The most abundant non-quinone compound was isopentyl eicosenoate. The relative abundance of quinone and non-quinone in the defensive fluid of U. transsilvanicus was 77% and 23%, respectively. The phylogenetic importance of the registered compounds is briefly discussed. Key words: Diplopoda, Julidae, Unciger transsilvanicus, GC-MS, defense secretion, quinones, p-cresol, phenol, isopentyl esters of saturated and unsaturated fatty acids from C14 to C20, isopentyl hexacosatetraenoate INTRODUCTION The millipede family Julidae includes more than 600 species and represents the dominant family of A wide variety of arthropod species has evolved Diplopoda in the western part of the Palaearctic re- chemical defense mechanisms that have contributed gion (Enghoff and Kime, 2011). -
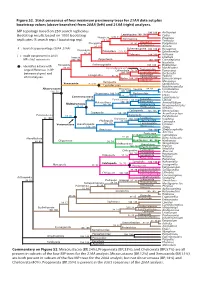
Figure S2. Strict Consensus of Four Maximum Parsimony Trees for 21AA Data Set Plus Bootstrap Values (Above Branches) from 20AA (Left) and 21AA (Right) Analyses
Figure S2. Strict consensus of four maximum parsimony trees for 21AA data set plus bootstrap values (above branches) from 20AA (left) and 21AA (right) analyses. MP topology based on 250 search replicates. 100 100 Antheraea Bootstrap results based on 1000 bootstrap Lepidoptera 100 100 Cydia Neoptera [23] 44 Dermaptera Prodoxus replicates (5 search reps / bootstrap rep). 68 66 Forficula Blattodea Pterygota 92 96 Periplaneta Orthoptera 96 97 Acheta # : bootstrap percentage (20AA 21AA) Ephemeroptera Dicondylia 100 100 Hexagenia Paleoptera [37] 52 Ephemerella 98 98 Odonata 100 100 Ischnura [ ] : node not present in 20AA Insecta Libellula MP strict consensus 100 100 Zygentoma 100 100 Ctenolepisma Nicoletia Archaeognatha : Identifies 6 taxa with Hexapoda 100 100 Pedetontus 89 86 Machiloides Entomobryomorpha Tomoceridae large differences in BP Collembola 70 76 Tomocerus between degen1 and 100 100 Entomobryidae Orchesella Entognatha 82 82 Poduridae Podura 20AA analyses. Diplura 100 100 Campodeidae Japygidae Eumesocampa Remipedia Metajapyx Xenocarida [31] 51 Speleonectes Cephalocarida Hutchinsoniella Altocrustacea Thoracica Sessilia 88 83 Semibalanus [14] 31 100 100 Chthamalus Thecostraca 100 100 Pedunculata Lepas Communostraca Rhizocephala Loxothylacus 96 89 Eumalacostraca 84 85 Eucarida Libinia Malacostraca 100 100 Peracarida Armadillidium Multicrustacea 100 100 Hoplocarida 69 84 Neogonodactylus Phyllocarida Nebalia Cyclopoida 100 100 Mesocyclops Copepoda 100 100 Acanthocyclops Pancrustacea 65 82 Calanoida Eurytemora 96 100 Diplostraca 100 100 -

Some Aspects of the Ecology of Millipedes (Diplopoda) Thesis
Some Aspects of the Ecology of Millipedes (Diplopoda) Thesis Presented in Partial Fulfillment of the Requirements for the Degree Master of Science in the Graduate School of The Ohio State University By Monica A. Farfan, B.S. Graduate Program in Evolution, Ecology, and Organismal Biology The Ohio State University 2010 Thesis Committee: Hans Klompen, Advisor John W. Wenzel Andrew Michel Copyright by Monica A. Farfan 2010 Abstract The focus of this thesis is the ecology of invasive millipedes (Diplopoda) in the family Julidae. This particular group of millipedes are thought to be introduced into North America from Europe and are now widely found in many urban, anthropogenic habitats in the U.S. Why are these animals such effective colonizers and why do they seem to be mostly present in anthropogenic habitats? In a review of the literature addressing the role of millipedes in nutrient cycling, the interactions of millipedes and communities of fungi and bacteria are discussed. The presence of millipedes stimulates fungal growth while fungal hyphae and bacteria positively effect feeding intensity and nutrient assimilation efficiency in millipedes. Millipedes may also utilize enzymes from these organisms. In a continuation of the study of the ecology of the family Julidae, a comparative study was completed on mites associated with millipedes in the family Julidae in eastern North America and the United Kingdom. The goals of this study were: 1. To establish what mites are present on these millipedes in North America 2. To see if this fauna is the same as in Europe 3. To examine host association patterns looking specifically for host or habitat specificity. -

Order CALLIPODIDA Manual Versión Española
Revista IDE@ - SEA, nº 25B (30-06-2015): 1–12. ISSN 2386-7183 1 Ibero Diversidad Entomológica @ccesible www.sea-entomologia.org/IDE@ Class: Diplopoda Order CALLIPODIDA Manual Versión española CLASS DIPLOPODA Order Callipodida Jörg Spelda Bavarian State Collection of Zoology Münchhausenstraße 21, 81247 Munich, Germany [email protected] 1. Brief characterization of the group and main diagnostic characters 1.1. Morphology The members of the order Callipodida are best recognized by their putative apomorphies: a divided hypoproct, divided anal valves, long extrusible tubular vulvae, and, as in all other helminthomorph milli- pede orders, a characteristic conformation of the male gonopods. As in Polydesmida, only the first leg pair of the 7th body ring is transformed into gonopods, which are retracted inside the body. Body rings are open ventrally and are not fused with the sternites, leaving the coxae of the legs free. Legs in the anterior half of the body carry coxal pouches. The small collum does not overlap the head. Callipodida are of uniformly cylindrical external appearance. The number of body rings is only sometimes fixed in species and usually exceeds 40. There are nine antennomeres, as the 2nd antennomere of other Diplopoda is subdivided (= antennomere 2 and 3 in Callipodida). The general struc- ture of the gnathochilarium is shared with the Chordeumatida and Polydesmida. Callipodida are said to be characterised by longitudinal crests, which gives the order the common name “crested millipedes”. Although crest are present in most species, some genera (e.g. Schizopetalum) lack a crest, while some Spirostreptida ( e.g. in Cambalopsidae, ‘Trachystreptini’) and some Julida (e.g.