Regulators of HIV-1 Reverse Transcription
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Introduction of Human Telomerase Reverse Transcriptase to Normal Human Fibroblasts Enhances DNA Repair Capacity
Vol. 10, 2551–2560, April 1, 2004 Clinical Cancer Research 2551 Introduction of Human Telomerase Reverse Transcriptase to Normal Human Fibroblasts Enhances DNA Repair Capacity Ki-Hyuk Shin,1 Mo K. Kang,1 Erica Dicterow,1 INTRODUCTION Ayako Kameta,1 Marcel A. Baluda,1 and Telomerase, which consists of the catalytic protein subunit, No-Hee Park1,2 human telomerase reverse transcriptase (hTERT), the RNA component of telomerase (hTR), and several associated pro- 1School of Dentistry and 2Jonsson Comprehensive Cancer Center, University of California, Los Angeles, California teins, has been primarily associated with maintaining the integ- rity of cellular DNA telomeres in normal cells (1, 2). Telomer- ase activity is correlated with the expression of hTERT, but not ABSTRACT with that of hTR (3, 4). Purpose: From numerous reports on proteins involved The involvement of DNA repair proteins in telomere main- in DNA repair and telomere maintenance that physically tenance has been well documented (5–8). In eukaryotic cells, associate with human telomerase reverse transcriptase nonhomologous end-joining requires a DNA ligase and the (hTERT), we inferred that hTERT/telomerase might play a DNA-activated protein kinase, which is recruited to the DNA role in DNA repair. We investigated this possibility in nor- ends by the DNA-binding protein Ku. Ku binds to hTERT mal human oral fibroblasts (NHOF) with and without ec- without the need for telomeric DNA or hTR (9), binds the topic expression of hTERT/telomerase. telomere repeat-binding proteins TRF1 (10) and TRF2 (11), and Experimental Design: To study the effect of hTERT/ is thought to regulate the access of telomerase to telomere DNA telomerase on DNA repair, we examined the mutation fre- ends (12, 13). -

Purification and Characterization of the Reverse Transcriptase Expressed In
Volume 270, number 1,2, 76-80 FEBS 08845 September 1990 Purification and characterization of the RNase H domain of HIV-l reverse transcriptase expressed in recombinant Escherichia coli S. Patricia Becerra’, G. Marius Clore2, Angela M. Gronenbornz, Anders R. Karlstr6m3, Stephen J. Stahl‘+, Samuel H. Wilson’ and Paul T. Wingfield 1Laboratory of Biochemistry, NCI, ZLaboratory of Chemical Physics, NIDDK, 3Laboratory of Biochemistry, NHLB and 4Protein Expression Laboratory, National Institutes of Health, Bethesda, MD 20892, USA Received 2 July 1990 The ribonuclease H (RNase H) domain of human immuno-deficiency virus (HIV-l) reverse transcriptase has been produced with the aim of provid- ing sufficient amounts of protein for biophysical studies. A plasmid vector is described which directs high level expression of the RNase H domain under the control of the I PL promoter. The domain corresponds to residues 427-560 of the 66 kDa reverse transcriptase. The protein was expressed in Escherichia cob’ and was purified using ion-exchange and size exclusion chromatography. The purified protein appears to be in a native-like homogeneous conformational state as determined by iH-NMR spectroscopy and circular dichroism measurements. HIV-protease treatment of the RNase H domain resulted in cleavage between Phe-440 and Tyr-441. HIV-l reverse transcriptase; HIV-l RNase H; HIV protease; Protein expression; Protein purification; Protein conformation 1. INTRODUCTION as a separate 15 kDa protein. The purified protein ap- pears suitable for structural studies by both NMR spec- Ribonuclease H (RNase H) activity resides in the C- troscopy and X-ray analysis, thereby providing an addi- terminal region of retroviral polymerases as shown by tional target in the search for therapeutic agents against deletion experiments [ 11, linker insertion [2] and point the spread of HIV infection. -

Rapidxfire Thermostable Reverse Transcriptase Application Note
Application note RapiDxFire Thermostable Reverse Transcriptase Introduction RNA target, which improves cDNA yield and A reverse transcriptase (RT) is a DNA subsequent detection sensitivity from difficult polymerase enzyme that synthesises cDNA RNA targets. Thermostability also is an from an RNA template. RTs are widely indicator of general enzyme stability for storage, used to study gene expression in cells or automation applications, and lyophilisation. tissues, in next-generation sequencing Processivity, which refers to the number of (NGS) applications, and in conjunction with nucleotides incorporated into cDNA during quantitative PCR (qPCR) or isothermal a single enzyme binding event, can affect amplification methods to detect and identify cDNA length, RT efficiency, and RT reaction RNAs that are of clinical or functional time. Native RT enzymes also have RNase H significance. endonuclease activity that will cleave the RNA from a DNA-RNA duplex and limit cDNA length. RT enzymes may differ in key characteristics Some RT variants also have been engineered that affect their performance in different to reduce RNase H activity to allow longer applications. Their thermostability allows cDNA products. cDNA synthesis to be performed at higher temperatures (above 50 °C), enabling them Commercially available RTs used in molecular to melt areas of secondary structure in the biology are generally derived from Moloney Application note RapiDxFire Thermostable Reverse Transcriptase murine leukaemia virus (MMLV) or avian Skeletal Muscle Total RNA (Thermo Fisher myeloblastosis virus (AMV), which have optimal Scientific Cat No. AM7982), RNA, MS2 (Roche activity at 37-42 °C. Cloned AMV RT and Cat No. 10165948001), Zika virus ATCC® engineered variants of AMV RT and MMLV VR-1843™ (ATCC Cat No. -

Multiple Cellular Proteins Interact with LEDGF/P75 Through a Conserved Unstructured Consensus Motif
ARTICLE Received 19 Jan 2015 | Accepted 1 Jul 2015 | Published 6 Aug 2015 DOI: 10.1038/ncomms8968 Multiple cellular proteins interact with LEDGF/p75 through a conserved unstructured consensus motif Petr Tesina1,2,3,*, Katerˇina Cˇerma´kova´4,*, Magdalena Horˇejsˇ´ı3, Katerˇina Procha´zkova´1, Milan Fa´bry3, Subhalakshmi Sharma4, Frauke Christ4, Jonas Demeulemeester4, Zeger Debyser4, Jan De Rijck4,**, Va´clav Veverka1,** & Pavlı´na Rˇeza´cˇova´1,3,** Lens epithelium-derived growth factor (LEDGF/p75) is an epigenetic reader and attractive therapeutic target involved in HIV integration and the development of mixed lineage leukaemia (MLL1) fusion-driven leukaemia. Besides HIV integrase and the MLL1-menin complex, LEDGF/p75 interacts with various cellular proteins via its integrase binding domain (IBD). Here we present structural characterization of IBD interactions with transcriptional repressor JPO2 and domesticated transposase PogZ, and show that the PogZ interaction is nearly identical to the interaction of LEDGF/p75 with MLL1. The interaction with the IBD is maintained by an intrinsically disordered IBD-binding motif (IBM) common to all known cellular partners of LEDGF/p75. In addition, based on IBM conservation, we identify and validate IWS1 as a novel LEDGF/p75 interaction partner. Our results also reveal how HIV integrase efficiently displaces cellular binding partners from LEDGF/p75. Finally, the similar binding modes of LEDGF/p75 interaction partners represent a new challenge for the development of selective interaction inhibitors. 1 Institute of Organic Chemistry and Biochemistry of the ASCR, v.v.i., Flemingovo nam. 2, 166 10 Prague, Czech Republic. 2 Department of Genetics and Microbiology, Faculty of Science, Charles University in Prague, Vinicna 5, 128 44 Prague, Czech Republic. -

Educator Materials Hands-On Activity HIV Reverse Transcription and AZT
Hands-on Activity HIV Reverse Transcription and AZT Educator Materials HIV REVERSE TRANSCRIPTION AND AZT OVERVIEW This hands-on activity is part of a series of activities and demonstrations focusing on various aspects of the human immunodeficiency virus (HIV) life cycle. In this activity, students will model how the anti-HIV drug AZT (azidothymidine) interferes with the process of viral replication and reduces the amount of virus in the Body. Students will first model reverse transcription, the process that results in the production of a double- stranded DNA copy of the HIV single-stranded RNA genome. (See introduction in the student document for a review of the process.) Using an actual HIV RNA sequence as a template, students will model the synthesis of a complementary strand of DNA By attaching nucleotides to one another. Then, students will demonstrate AZT’s effect on this process. They will suBstitute AZT in place of thymidine. AZT lacks a hydroxyl (OH) group that is necessary for suBsequent nucleotides to attach. When AZT is incorporated in a growing DNA sequence it prevents further nucleotides from Being added, thereby blocking the production of HIV DNA. Following this activity, it may Be helpful for students to complete the HIV integration activity. LEARNING OBJECTIVES Students will Be able to: • Code DNA from RNA. • Demonstrate the process of reverse transcription. • Understand the role of the enzyme reverse transcriptase in the formation of viral DNA. • Review the chemical structures of nucleic acids and identify structural -

HIV-1 Rev Downregulates Tat Expression and Viral Replication Via Modulation of NAD(P)H:Quinine Oxidoreductase 1 (NQO1)
ARTICLE Received 25 Jul 2014 | Accepted 22 Apr 2015 | Published 10 Jun 2015 DOI: 10.1038/ncomms8244 HIV-1 Rev downregulates Tat expression and viral replication via modulation of NAD(P)H:quinine oxidoreductase 1 (NQO1) Sneh Lata1,*, Amjad Ali2,*, Vikas Sood1,2,w, Rameez Raja2 & Akhil C. Banerjea2 HIV-1 gene expression and replication largely depend on the regulatory proteins Tat and Rev, but it is unclear how the intracellular levels of these viral proteins are regulated after infection. Here we report that HIV-1 Rev causes specific degradation of cytoplasmic Tat, which results in inhibition of HIV-1 replication. The nuclear export signal (NES) region of Rev is crucial for this activity but is not involved in direct interactions with Tat. Rev reduces the levels of ubiquitinated forms of Tat, which have previously been reported to be important for its transcriptional properties. Tat is stabilized in the presence of NAD(P)H:quinine oxidoreductase 1 (NQO1), and potent degradation of Tat is induced by dicoumarol, an NQO1 inhibitor. Furthermore, Rev causes specific reduction in the levels of endogenous NQO1. Thus, we propose that Rev is able to induce degradation of Tat indirectly by downregulating NQO1 levels. Our findings have implications in HIV-1 gene expression and latency. 1 Department of Microbiology, University College of Medical Sciences and Guru Teg Bahadur Hospital, Delhi 110095, India. 2 Laboratory of Virology, National Institute of Immunology, New Delhi 110067, India. * These authors contributed equally to this work. w Present address: Translational Health Science and Technology Institute, Faridabad, Haryana 121004, India. Correspondence and requests for materials should be addressed to A.C.B. -

Viroporins: Structures and Functions Beyond Cell Membrane Permeabilization
Editorial Viroporins: Structures and Functions beyond Cell Membrane Permeabilization José Luis Nieva 1,* and Luis Carrasco 2,* Received: 17 September 2015 ; Accepted: 21 September 2015 ; Published: 29 September 2015 Academic Editor: Eric O. Freed 1 Biophysics Unit (CSIC, UPV/EHU) and Biochemistry and Molecular Biology Department, University of the Basque Country (UPV/EHU), P.O. Box 644, 48080 Bilbao, Spain 2 Centro de Biología Molecular Severo Ochoa (CSIC, UAM), c/Nicolás Cabrera, 1, Universidad Autónoma de Madrid, Cantoblanco, 28049 Madrid, Spain * Correspondence: [email protected] (J.L.N.); [email protected] (L.C.); Tel.: +34-94-601-3353 (J.L.N.); +34-91-497-8450 (L.C.) Viroporins represent an interesting group of viral proteins that exhibit two sets of functions. First, they participate in several viral processes that are necessary for efficient production of virus progeny. Besides, viroporins interfere with a number of cellular functions, thus contributing to viral cytopathogenicity. Twenty years have elapsed from the first review on viroporins [1]; since then several reviews have covered the advances on viroporin structure and functioning [2–8]. This Special Issue updates and revises new emerging roles of viroporins, highlighting their potential use as antiviral targets and in vaccine development. Viroporin structure. Viroporins are usually short proteins with at least one hydrophobic amphipathic helix. Homo-oligomerization is achieved by helix–helix interactions in membranes rendering higher order structures, forming aqueous pores. Progress in viroporin structures during the last 2–3 years has in some instances provided a detailed knowledge of their functional architecture, including the fine definition of binding sites for effective inhibitors. -
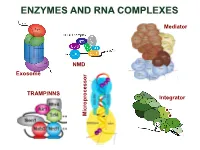
Enzymes and Rna Complexes
ENZYMES AND RNA COMPLEXES Mediator NMD Exosome NMD TRAMP/NNS Integrator Microprocessor RNA PROCESSING and DECAY machinery: RNases Protein Function Characteristics Exonucleases 5’ 3’ Xrn1 cytoplasmic, mRNA degradation processsive Rat1 nuclear, pre-rRNA, sn/snoRNA, pre-mRNA processing and degradation Rrp17/hNol12 nuclear, pre-rRNA processing Exosome 3’ 5’ multisubunit exo/endo complex subunits organized as in bacterial PNPase Rrp44/Dis3 catalytic subunit Exo/PIN domains, processsive Rrp4, Rrp40 pre-rRNA, sn/snoRNA processing, mRNA degradation Rrp41-43, 45-46 participates in NMD, ARE-dependent, non-stop decay Mtr3, Ski4 Mtr4 nuclear helicase cofactor DEAD box Rrp6 (Rrp47) nuclear exonuclease ( Rrp6 BP, cofactor) RNAse D homolog, processsive Ski2,3,7,8 cytoplasmic exosome cofactors. SKI complex helicase, GTPase Other 3’ 5’ Rex1-4 3’-5’ exonucleases, rRNA, snoRNA, tRNA processing RNase D homolog DXO 3’-5’ exonuclease in addition to decapping mtEXO 3’ 5’ mitochondrial degradosome RNA degradation in yeast Suv3/ Dss1 helicase/ 3’-5’ exonuclease DExH box/ RNase II homolog Deadenylation Ccr4/NOT/Pop2 major deadenylase complex (Ccr, Caf, Pop, Not proteins) Ccr4- Mg2+ dependent endonuclease Pan2p/Pan3 additional deadenylases (poliA tail length) RNase D homolog, poly(A) specific nuclease PARN mammalian deadenylase RNase D homolog, poly(A) specific nuclease Endonucleases RNase III -Rnt1 pre-rRNA, sn/snoRNA processing, mRNA degradation dsRNA specific -Dicer, Drosha siRNA/miRNA biogenesis, functions in RNAi PAZ, RNA BD, RNase III domains Ago2 Slicer -

Allosteric Integrase Inhibitor Potency Is Determined Through the Inhibition of HIV-1 Particle Maturation
Allosteric integrase inhibitor potency is determined through the inhibition of HIV-1 particle maturation Kellie A. Juradoa, Hao Wanga, Alison Slaughterb, Lei Fengb, Jacques J. Kesslb, Yasuhiro Koha, Weifeng Wanga, Allison Ballandras-Colasa, Pratiq A. Patelc, James R. Fuchsc, Mamuka Kvaratskheliab, and Alan Engelmana,1 aDepartment of Cancer Immunology and AIDS, Dana-Farber Cancer Institute and Department of Medicine, Harvard Medical School, Boston, MA 02215; and bCenter for Retrovirus Research and Comprehensive Cancer Center and cDivision of Medicinal Chemistry and Pharmacognosy, College of Pharmacy, The Ohio State University, Columbus, OH 43210 Edited by Alan R. Rein, National Cancer Institute, Frederick, MD, and accepted by the Editorial Board April 1, 2013 (received for review January 14, 2013) Integration is essential for HIV-1 replication, and the viral integrase HIV-1 preferentially integrates along the bodies of active genes (IN) protein is an important therapeutic target. Allosteric IN inhib- (6), a trait that is largely attributable to an interaction between itors (ALLINIs) that engage the IN dimer interface at the binding site IN and the host protein lens epithelium-derived growth factor for the host protein lens epithelium-derived growth factor (LEDGF)/ (LEDGF)/transcriptional coactivator p75 (reviewed in refs. 7 and transcriptional coactivator p75 are an emerging class of small mole- 8). LEDGF/p75 functions as a bimodal tether during integration: cule antagonists. Consistent with the inhibition of a multivalent drug elements within its N-terminal region confer constitutive binding to target, ALLINIs display steep antiviral dose–response curves ex vivo. chromatin, whereas a downstream IN-binding domain (IBD) binds ALLINIs multimerize IN protein and concordantly block its assembly lentiviral IN proteins (9, 10). -

The Primer Binding Site on the RNA Genome of Human and Simian Immunodeficiency Viruses Is Flanked by an Upstream Hairpin Structure Benjamin Berkhout
1997 Oxford University Press Nucleic Acids Research, 1997, Vol. 25, No. 20 4013–4017 The primer binding site on the RNA genome of human and simian immunodeficiency viruses is flanked by an upstream hairpin structure Benjamin Berkhout Department of Human Retrovirology, Academic Medical Center, University of Amsterdam, PO Box 22700, 1100 DE Amsterdam, The Netherlands Received July 16, 1997; Revised and Accepted August 26, 1997 ABSTRACT PBS site. Such additional contacts were originally proposed for Rous sarcoma virus (2–4), but similar interactions have been Reverse transcription of retroviral genomes is primed reported for human immunodeficiency virus type 1 (HIV-1) (5), by a tRNA molecule that anneals to an 18 nt primer HIV-2 (6) and several retrotransposon elements (7–9). Consistent binding site (PBS) on the viral RNA genome. Additional with the idea that regions of the viral genome outside the PBS base pair interactions between the tRNA primer and sequence participate in selective tRNA primer usage is the the viral RNA have been proposed. In particular, base observation that retroviruses mutated in the PBS are genetically pairing was proposed between the anticodon loop of unstable, as they revert to the wild-type PBS sequence after a few tRNALys3 and the ‘A-rich’ loop of a hairpin located passages in cell culture (10–13). On the other hand, reasonable immediately upstream of the PBS site in HIV-1 RNA. In transduction efficiencies were obtained with murine leukemia order to judge the importance of this sequence/structure virus vectors that use an unnatural or genetically engineered motif, we performed an extensive phylogenetic tRNA primer (14,15). -

Human Retroviruses in the Second Decade: a Personal Perspective
© 1995 Nature Publishing Group http://www.nature.com/naturemedicine • REVIEW Human retroviruses in the second decade: A personal perspective Human retroviruses have developed novel strategies for their propagation and survival. A consequence of their success has been the induction of an extraordinarily diverse set of human dlst!ases, including AIDS, cancers and neurological and Inflammatory disorders. Early research focused on their characterization, linkage to these dlst!ases, and the mechanisms Involved. Research should now aim at the eradication of human retroviruses and on treatment of infected people. Retroviruses are transmitted either geneti- .................... ···.. .... ·. ..... .. discovered". Though its characteristics are cally (endogenous form) or as infectious ROBERT C. GALLO strikingly similar to HTLV-1, HTLV-II is not agents (exogenous form)'·'. As do many so clearly linked to human disease. It is cu other animal species, humans have both forms ..... In general, rious that HTLV-11 is endemic in some American Indians and endogenous retroviruses are evolutionary relics of old infec more prevalent in drug addicts than HTLV-I"·'•. tions and are not known to cause disease. The DNA of many HIV-1 is also most prevalent in equatorial Africa, but in con species, including humans, harbours multiple copies of differ trast to HTLV the demography of the HIV epidemic is still in ent retroviral proviruses. The human endogenous proviral flux, and the virus is new to most of the world. The number of sequences are virtually all defective, and comprise about one infected people worldwide is now estimated to be about 17 mil percent of the human genome, though R. Kurth's group in lion and is predicted to reach 30 to SO million by the year 2000. -

Ribonuclease A
Chem. Rev. 1998, 98, 1045−1065 1045 Ribonuclease A Ronald T. Raines Departments of Biochemistry and Chemistry, University of WisconsinsMadison, Madison, Wisconsin 53706 Received October 10, 1997 (Revised Manuscript Received January 12, 1998) Contents I. Introduction 1045 II. Heterologous Production 1046 III. Structure 1046 IV. Folding and Stability 1047 A. Disulfide Bond Formation 1047 B. Prolyl Peptide Bond Isomerization 1048 V. RNA Binding 1048 A. Subsites 1048 B. Substrate Specificity 1049 C. One-Dimensional Diffusion 1049 D. Processive Catalysis 1050 VI. Substrates 1050 VII. Inhibitors 1051 Ronald T. Raines was born in 1958 in Montclair, NJ. He received Sc.B. VIII. Reaction Mechanism 1052 degrees in chemistry and biology from the Massachusetts Institute of A. His12 and His119 1053 Technology. At M.I.T., he worked with Christopher T. Walsh to reveal the reaction mechanisms of pyridoxal 5′-phosphate-dependent enzymes. B. Lys41 1054 Raines was a National Institutes of Health predoctoral fellow in the C. Asp121 1055 chemistry department at Harvard University. There, he worked with D. Gln11 1056 Jeremy R. Knowles to elucidate the reaction energetics of triosephosphate IX. Reaction Energetics 1056 isomerase. Raines was a Helen Hay Whitney postdoctoral fellow in the biochemistry and biophysics department at the University of California, A. Transphosphorylation versus Hydrolysis 1056 San Francisco. At U.C.S.F., he worked with William J. Rutter to clone, B. Rate Enhancement 1057 express, and mutate the cDNA that codes for ribonuclease A. Raines X. Ribonuclease S 1058 then joined the faculty of the biochemistry department at the University s A. S-Protein−S-Peptide Interaction 1058 of Wisconin Madison, where he is now associate professor of biochem- istry and chemistry.