Late Holocene Sea-Level Change Around Newfoundland Julia Daly
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
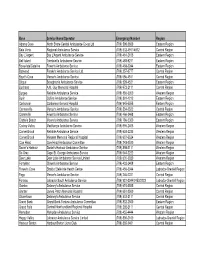
Revised Emergency Contact #S for Road Ambulance Operators
Base Service Name/Operator Emergency Number Region Adams Cove North Shore Central Ambulance Co-op Ltd (709) 598-2600 Eastern Region Baie Verte Regional Ambulance Service (709) 532-4911/4912 Central Region Bay L'Argent Bay L'Argent Ambulance Service (709) 461-2105 Eastern Region Bell Island Tremblett's Ambulance Service (709) 488-9211 Eastern Region Bonavista/Catalina Fewer's Ambulance Service (709) 468-2244 Eastern Region Botwood Freake's Ambulance Service Ltd. (709) 257-3777 Central Region Boyd's Cove Mercer's Ambulance Service (709) 656-4511 Central Region Brigus Broughton's Ambulance Service (709) 528-4521 Eastern Region Buchans A.M. Guy Memorial Hospital (709) 672-2111 Central Region Burgeo Reliable Ambulance Service (709) 886-3350 Western Region Burin Collins Ambulance Service (709) 891-1212 Eastern Region Carbonear Carbonear General Hospital (709) 945-5555 Eastern Region Carmanville Mercer's Ambulance Service (709) 534-2522 Central Region Clarenville Fewer's Ambulance Service (709) 466-3468 Eastern Region Clarke's Beach Moore's Ambulance Service (709) 786-5300 Eastern Region Codroy Valley MacKenzie Ambulance Service (709) 695-2405 Western Region Corner Brook Reliable Ambulance Service (709) 634-2235 Western Region Corner Brook Western Memorial Regional Hospital (709) 637-5524 Western Region Cow Head Cow Head Ambulance Committee (709) 243-2520 Western Region Daniel's Harbour Daniel's Harbour Ambulance Service (709) 898-2111 Western Region De Grau Cape St. George Ambulance Service (709) 644-2222 Western Region Deer Lake Deer Lake Ambulance -
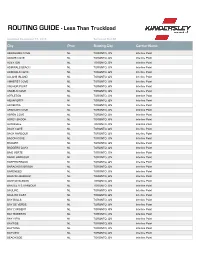
ROUTING GUIDE - Less Than Truckload
ROUTING GUIDE - Less Than Truckload Updated December 17, 2019 Serviced Out Of City Prov Routing City Carrier Name ABRAHAMS COVE NL TORONTO, ON Interline Point ADAMS COVE NL TORONTO, ON Interline Point ADEYTON NL TORONTO, ON Interline Point ADMIRALS BEACH NL TORONTO, ON Interline Point ADMIRALS COVE NL TORONTO, ON Interline Point ALLANS ISLAND NL TORONTO, ON Interline Point AMHERST COVE NL TORONTO, ON Interline Point ANCHOR POINT NL TORONTO, ON Interline Point ANGELS COVE NL TORONTO, ON Interline Point APPLETON NL TORONTO, ON Interline Point AQUAFORTE NL TORONTO, ON Interline Point ARGENTIA NL TORONTO, ON Interline Point ARNOLDS COVE NL TORONTO, ON Interline Point ASPEN COVE NL TORONTO, ON Interline Point ASPEY BROOK NL TORONTO, ON Interline Point AVONDALE NL TORONTO, ON Interline Point BACK COVE NL TORONTO, ON Interline Point BACK HARBOUR NL TORONTO, ON Interline Point BACON COVE NL TORONTO, ON Interline Point BADGER NL TORONTO, ON Interline Point BADGERS QUAY NL TORONTO, ON Interline Point BAIE VERTE NL TORONTO, ON Interline Point BAINE HARBOUR NL TORONTO, ON Interline Point BAKERS BROOK NL TORONTO, ON Interline Point BARACHOIS BROOK NL TORONTO, ON Interline Point BARENEED NL TORONTO, ON Interline Point BARR'D HARBOUR NL TORONTO, ON Interline Point BARR'D ISLANDS NL TORONTO, ON Interline Point BARTLETTS HARBOUR NL TORONTO, ON Interline Point BAULINE NL TORONTO, ON Interline Point BAULINE EAST NL TORONTO, ON Interline Point BAY BULLS NL TORONTO, ON Interline Point BAY DE VERDE NL TORONTO, ON Interline Point BAY L'ARGENT NL TORONTO, ON -

Hearing Aid Providers (Including Audiologists)
Hearing Aid Providers (including Audiologists) Beltone Audiology and Hearing Clinic Inc. 16 Pinsent Drive Grand Falls-Windsor, NL A2A 2R6 1-709-489-8500 (Grand Falls-Windsor) 1-866-489-8500 (Toll free) 1-709-489-8497 (Fax) [email protected] Audiologist: Jody Strickland Hearing Aid Practitioners: Joanne Hunter, Jodine Reid Satellite Locations: Baie Verte, Burgeo, Cow Head, Flower’s Cove, Harbour Breton, Port Saunders, Springdale, Stephenville, St. Albans, St. Anthony ________________________________ Beltone Audiology and Hearing Clinic Inc. 3 Herald Avenue Corner Brook, NL A2H 4B8 1-709-639-8501 (Corner Brook) 1-866-489-8500 (Toll free) 1-709-639-8502 (Fax) [email protected] Audiologist: Jody Strickland Hearing Aid Practitioner: Jason Gedge Satellite Locations: Baie Verte, Burgeo, Cow Head, Flower’s Cove, Harbour Breton, Port Saunders, Springdale, Stephenville, St. Albans, St. Anthony ________________________________ Beltone Hearing Service 3 Paton Street St. John’s, NL A1B 4S8 1-709-726-8083 (St. John’s) 1-800-563-8083 (Toll free) 1-709-726-8111 (Fax) [email protected] www.beltone.nl.ca Audiologists: Brittany Green Hearing Aid Practitioners: Mike Edwards, David King, Kim King, Joe Lynch, Lori Mercer Satellite Locations: Bay Roberts, Bonavista, Carbonear, Clarenville, Gander, Grand Bank, Happy Valley-Goose Bay, Labrador City, Lewisporte, Marystown, New-Wes- Valley, Twillingate ________________________________ Exploits Hearing Aid Centre 9 Pinsent Drive Grand Falls-Windsor, NL A2A 2S8 1-709-489-8900 (Grand Falls-Windsor) 1-800-563-8901 (Toll free) 1-709-489-9006 (Fax) [email protected] www.exploitshearing.ca Hearing Aid Practitioners: Dianne Earle, William Earle, Toby Penney ________________________________ Maico Hearing Service 84 Thorburn Road St. -

National Librarian Visits the Atlantic Region
• e l The Atlantic Provinces Library Association Volume 63, Number 4 ISSN 0001-2203 January / February 2000 National Librarian Visits the Atlantic Region The day's library issues forum took place, between Left to right: Jocelyne Thompson, New Brunswick Provincial Librarian; Roch Carrier, National Librarian; John Teshcy, 12:00 and 2:00 at the Lord Dalhousie Room, on University Librarian, UNB; Sara Lochhead, University Librarian, Dalhousie University campus, hosted hy the Dalhousie Mount Allison University School of Information Studies, and moderated by the schools director Bertrum MacDonald. The forum was NOVA SCOTIA: Tuesday November 30,1999 well attended by over 60 people. Fourteen speakers gave For Roch Carrier's visit, arrangements in Halifax brief presentations. They represented Nova Scotia were ably coordinated by the staff of the Nova Scotia library associations and agencies and a wide range of Provincial Library, with assistance and support from a library sectors, including school, university, government number of individuals and organizations. Roch Carrier and law libraries, as well as library students, public and began the day with a breakfast with staff of the Nova other special libraries. A wide range of library initiatives Scotia Provincial Library. He then toured the Terrence and pressing issues were discussed. Themes which came Bay Elementary School Library. Next was a visit to the up repeatedly, were the need for resource sharing ven library of the John A. MacDonald High school, and a tures, and centralized leadership to find innovative ways meeting with students there. Then came a visit to the to address the growing resource shortages our institu Halifax Regional Public Library, Spryfield Branch. -

Rental Housing Portfolio March 2021.Xlsx
Rental Housing Portfolio Profile by Region - AVALON - March 31, 2021 NL Affordable Housing Partner Rent Federal Community Community Housing Approved Units Managed Co-op Supplement Portfolio Total Total Housing Private Sector Non Profit Adams Cove 1 1 Arnold's Cove 29 10 39 Avondale 3 3 Bareneed 1 1 Bay Bulls 1 1 10 12 Bay Roberts 4 15 19 Bay de Verde 1 1 Bell Island 90 10 16 116 Branch 1 1 Brigus 5 5 Brownsdale 1 1 Bryants Cove 1 1 Butlerville 8 8 Carbonear 26 4 31 10 28 99 Chapel Cove 1 1 Clarke's Beach 14 24 38 Colinet 2 2 Colliers 3 3 Come by Chance 3 3 Conception Bay South 36 8 14 3 16 77 Conception Harbour 8 8 Cupids 8 8 Cupids Crossing 1 1 Dildo 1 1 Dunville 11 1 12 Ferryland 6 6 Fox Harbour 1 1 Freshwater, P. Bay 8 8 Gaskiers 2 2 Rental Housing Portfolio Profile by Region - AVALON - March 31, 2021 NL Affordable Housing Partner Rent Federal Community Community Housing Approved Units Managed Co-op Supplement Portfolio Total Total Housing Goobies 2 2 Goulds 8 4 12 Green's Harbour 2 2 Hant's Harbour 0 Harbour Grace 14 2 6 22 Harbour Main 1 1 Heart's Content 2 2 Heart's Delight 3 12 15 Heart's Desire 2 2 Holyrood 13 38 51 Islingston 2 2 Jerseyside 4 4 Kelligrews 24 24 Kilbride 1 24 25 Lower Island Cove 1 1 Makinsons 2 1 3 Marysvale 4 4 Mount Carmel-Mitchell's Brook 2 2 Mount Pearl 208 52 18 10 24 28 220 560 New Harbour 1 10 11 New Perlican 0 Norman's Cove-Long Cove 5 12 17 North River 4 1 5 O'Donnels 2 2 Ochre Pit Cove 1 1 Old Perlican 1 8 9 Paradise 4 14 4 22 Placentia 28 2 6 40 76 Point Lance 0 Port de Grave 0 Rental Housing Portfolio Profile by Region - AVALON - March 31, 2021 NL Affordable Housing Partner Rent Federal Community Community Housing Approved Units Managed Co-op Supplement Portfolio Total Total Housing Portugal Cove/ St. -

8 Day Newfoundland Discovery
Tour Code NFLD 8 Day Newfoundland Discovery 8 days Created on: 29 Sep, 2021 Price Day 1: Arrive in Corner Brook, NL Arrive in Corner Brook in Newfoundland, Canada's most easterly province and also known as "The Rock". The town of Corner Brook is located on the west side of the island and renowned for its world-famous salmon bearing river, the Humber. Overnight: Corner Brook Day 2: Corner Brook - Cow Head, NL Today we explore incredible Gros Morne National Park, with its towering inland fjords and walking trails. We will learn about the geology of the area at the Gros Morne Visitor Centre and experience it first hand with a scenic walk through the Tablelands area discovering the unique flora and fauna native to the region. Moose, caribou, waterfalls and dozens of unforgettable photographic scenes add to this UNESCO World Heritage Destination. Next we visit Lobster Cove Lighthouse in Rocky Harbour and enjoy a boat cruise of Bonne Bay and participate in an age-old tradition - the "Screech In" ceremony, featuring live traditional music, a cod fish, and a taste of the famous Newfoundland Screech. Following the boat cruise, we continue up the Northern Peninsula of Newfoundland to the town of Cow Head. Overnight: Cow Head Included Meal(s): Breakfast, Lunch and Dinner Day 3: Cow Head - St. Anthony, NL This morning we travel the well-known Viking Trail Route to the most northerly tip of Newfoundland, to the L'Anse aux Meadows National Historic Site. We will see where the Vikings, the first Europeans to reach the New World, landed and established a settlement. -

2020/21 Fish Processors -Licences Expire March 31, 2021
2020/21 Fish Processors -Licences Expire March 31, 2021 Location of Postal Company Phone COMPANY NAME Processing Plant License Type Species Licensed for Head Office Community Contact Head Office Address P.O. Box Code Number 3 T's Limited Woody Point Primary Crab, Snow Woody Point Todd Young P.O. Box 71 A0K 1P0 709-453-2479 Seal Groundfish, All Species Lumpfish Pelagics, All Species Lobster 54417 Newfoundland and Labrador Inc. Harbour Breton Aquaculture Salmonids (Aquaculture) Corner Brook Bill Barry 415 Griffin Drive A2H 3E9 709-785-7387 Allen's Fisheries Limited Benoit's Cove Primary Crab, Snow Benoit's Cove Bill Barry A0L 1A0 709-785-7387 Pelagics, All Species Lumpfish Mussels (Aquaculture) Lobster Groundfish, All Species Oyster Aqua Crab Producers Inc. Aquaforte Primary Scallop Harbour Grace Joseph George P.O. Box 1986 A0A 2M0 709-596-7186 Crab, Snow Atlantic Treasure Seafoods Limited Bay Roberts Secondary Salmonids (Aquaculture) Bay Roberts David Russell P.O. Box 698 A0A 1G0 709-786-6712 Pelagics, All Species Seal Groundfish, All Species Avalon Ocean Products Incorporated Fair Haven Primary Lumpfish Arnold's Cove Mike Phillpott P.O. Box 40 A0B 1A0 709-463-8539 Groundfish, All Species Lobster Pelagics, All Species Scallop Barry Group Inc. Corner Brook Primary Pelagics, All Species Corner Brook Bill Barry 415 Griffin Drive A2H 3E9 709-785-7387 Groundfish, All Species Witless Bay Primary Lumpfish Crab, Snow Groundfish, All Species Cox's Cove Primary Lobster Groundfish, All Species Pelagics, All Species Dover Primary Pelagics, All Species Groundfish, All Species Salmonids (Aquaculture) Bay Roberts Seafoods Limited Bay Roberts Primary Lumpfish Bay Roberts David Russell P.O. -

2010-11 Annual Report
Annual Report 2010‐11 Submitted by The Provincial Information and Library Resources Board BOARD STRUCTURES Provincial Information and Library Resources Board The Provincial Information and Library Resources Board (PILRB) is an independent organization established by the Government of Newfoundland and Labrador, under authority of the Public Libraries Act, to oversee the operation of the public library services in the province now commonly referred to as the Newfoundland and Labrador Public Libraries (NLPL). The organization has existed, in some form, since 1935. The PILRB is a provincial board comprised of representatives and alternates of regional library boards and appointees of the Lieutenant‐Governor in Council. The provincial board has not less than 10 and not more than 15 members which include: (a) a representative from each regional library board appointed by that board; (b) the chairperson of the St. John's Library Board appointed by that board; and (c) up to six other members appointed by the Lieutenant‐ Governor in Council. The current board members, as of March 31, 2011, can be viewed in Appendix 1. Regional and Local Library Boards The PILRB currently operates 96 public libraries across the province. Each local library is operated by a local library board consisting of five to nine members with the exception of the three libraries in St. John’s, which operate under the St. John’s Library Board. A representative of each local library board is appointed to a regional library board, which assists the provincial board to ensure services and programs are consistent throughout the different regions of the province and aids in the development and implementation of policies. -
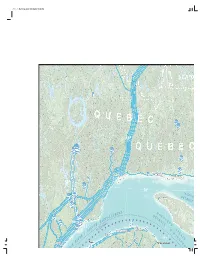
Q U E B C Q U É
(1,1) -1- Maritimes.indd 2020-06-04 10:30 AM U M 52°52°2 Q U E B C ManicouaganM anii couaouagag -70°-70-7700 ° -66°--66666°6 ° Q U É B E HavreHavravvrere de-Mingande-MMingani g Saint-PierreSainSSaSaiSaint i tin Pierret-Pit-Pierre PierrePierreerree 50° J A C Q U E D É S -Menier T R Port-MenierPortPoP rt O I T A N T Î I C L E O H S T T O N D I R E N D É G ’ A A U T U N - L R O E T I S N T I D I L A I T O C O S c S E b e S V u é D T -68° U Q ' H R L E t o O A F N I / Cap-Chatapap-ChatChatCh G T -64° U R E E Rivière-au-RenardR D I V R O E C N E (2,1) -1- Maritimes.indd 2020-06-04 10:30 AM U ND ND AND LAB TE E-N R DOR EUVE-ET- L RADOR e 52°52° 52° E L I S E L E I S L L E E L N B L E F B O E D I T L'AnseL'AnL''A see auau LoupLoL up A I T R O T R S T É D St. Anthony -62°-62626 ° Forresters Point E -58° -54° Port au Choix Englee AguanishAguaAAguang nishnisshh y a B P e Q U it E 50° h S W - C I T A R 5 D T I Cow Head 50° Seal Cove E E R J A C S ay A Q T e B U R Dam E A re ame S I ot re-D Fogo - C T N Not A de R ie Twillingate T Ba I S I E C L R Hampden O A Trout River Springdale S N T I D Lewisporte Deer Lake -60° e k Grand Falls- Corner Brook a Windsor Gander L r R i v e d s n n i t Bo a l o r Buchans p ie G E x Ba -56° NEWFOUNDLAND Glovertown AND LABRADOR (3,1) -1- Maritimes.indd 2020-06-04 10:30 AM Energy production and transmission IN ATLANTIC CANADA Production et transport de l’énergie AU50°50° CANADA ATLANTIQUE scale in kilometres -52°0 50 100 150 200 250 Bay a ist ista av nav on Bo B de e ai B Bonavista G U E Rivière-au-RenardR D O I fr R om (1,2) -1- Maritimes.indd 2020-06-04 10:30 AM GaspéGaspassppépé fr E ov om C er N N se G A S P É S I E as E E L A R Mont-ntt U L E D W JoliJoolili N I N S Percé A P É L . -

Tuckamore a Visitor's Guide BONNE BAY: D
Tuckamore A Visitor's Guide BONNE BAY: D. WILSON 1 Contact Information Gros Morne National Park of Canada P.O. Box 130, Table of Contents Rocky Harbour, NL A0K 4N0 2 World Heritage [email protected] www.pc.gc.ca/grosmorne 4 Unforgettable Adventures (709) 458-2417 6 Aquatic Encounters Campground Welcome 7 Western Brook Pond Reservations www.reservations. to Gros Morne National Park 8 Take a Hike parkscanada.gc.ca 10 Experience Proper 1-877-737-3783 / (TTY 1-866-787-6221) Awesome 12 Scenic Drives Swimming Pool (709) 458-2350 13 Conservation Boat Tours 14 Trail Guide/Map Western Brook Pond 16 Experience Gros Morne (709) 458-2016 / 1-888-458-2016 18 Camping Ferry Information/ 20 Wildlife Reservations I was born and raised in Norris Point, a small community in the middle of in this region (Gros Morne and L’Anse aux Meadows are the other two). 22 History and Heritage Marine Atlantic Gros Morne National Park. I’ve lived in a National Park for most of my life, In addition to the sites operated by Parks Canada Agency, there are many 1-800-341-7981 23 Communities and have spent my entire career working for Parks Canada. I have been very more historic sites, houses and museums managed by communities and Strait of Belle Isle lucky to have worked in places like Banff in Alberta and the Niagara National associations throughout western Newfoundland and Labrador. 24 Cultural Crossroads (St. Barbe) 1-866-535-2567 Historic Sites in Niagara on the Lake, Ontario. After an absence of more than 26 Port Au Choix five years, I am pleased to have had the opportunity to come home, and to On behalf of our passionate Parks Canada staff who you will meet along the 27 L’Anse Aux Meadows Bus and Taxi Services Martin’s Transportation welcome you all here as Superintendent. -

~Nalla~R~C (CANADA a GEOLOGY FIELD "GUIDE to SELECTED SITES in NEWFOUNDLAND, NOVA SCOTIA
D~s)COVER~NGROCK~~ ~j!NERAl~ ~NfO)FOs)S~l5) ~NAllA~r~C (CANADA A GEOLOGY FIELD "GUIDE TO SELECTED SITES IN NEWFOUNDLAND, NOVA SCOTIA, PRINCE EDV\JARDISLAND7 AND NEW BRUNSWICK 7_".-- ~ _. ...._ .•-- ~.- Peter Wallace. Editor Atlantic Geoscience Society Department of Earth Sciences La Societe G60scientifique Dalhousie University de L'Atlantique Halifax, Nova Scotia AGS Special Publication 14 • DISCOVERING ROCKS, MINERALS AND FOSSILS IN ATLANTIC CANADA A Geology Field Guide to Selected Sites in Newfoundland, Nova Scotia, Prince Edward Island and New Brunswick • Peter Wallace, editor Department of Earth Sciences Dalhousie University, Halifax, Nova Scotia Atlantic Geoscience Society La Societe Geoscientifique de L'Atlantique • AGS Special Publication • @ 1998 Atlantic Geoscience Society Department of Earth Sciences Dalhousie University 1236 Henry Street, Halifax Nova Scotia, Canada B3H3J5 This book was produced with help from The Canadian Geological Foundation, The Department of Earth Sciences, Dalhousie University, and The Atlantic Geoscience Society. ISBN 0-9696009-9-2 AGS Special Publication Number . 14.. I invite you to join the Atlantic Geoscience Society, write clo The Department of Earth Sciences, Dalhousie University (see above) Cover Photo Cape Split looking west into the Minas Channel, Nova Scotia. The split is caused by erosion along North-South faults cutting the Triassic-Jurassic-aged North Mountain Basalt and is the terminal point of a favoured hike of geologists and non-geologists alike. Photo courtesy of Rob • Fensome, Biostratigrapher, -

Tap Water Quality for Public Water Supplies in Newfoundland And
Water Resources Tap Water Quality for Public Water Supplies in Newfoundland Management Division and Labrador - Additional Parameters Community Name Serviced Area Source Name Sample Date Strontium Nitrate Nitrite TOC Units mg/L mg/L mg/L mg/L Guidelines for Canadian Drinking Water Quality 7 10 1 Anchor Point Anchor Point Well Cove Brook Feb 25, 2020 0.03 0.10 LTD 8.30 Appleton Appleton (+Glenwood) Gander Lake (The Outflow) Feb 03, 2020 0.01 0.10 LTD 6.80 Aquaforte Aquaforte Davies Pond Feb 05, 2020 0.01 LTD LTD 10.20 Arnold's Cove Arnold's Cove Steve's Pond (2 Intakes) Feb 27, 2020 0.01 LTD LTD 6.00 Avondale Avondale Lee's Pond Feb 18, 2020 0.01 LTD LTD 4.20 Baie Verte Baie Verte Southern Arm Pond Feb 12, 2020 0.01 LTD LTD 7.40 Baine Harbour Baine Harbour Baine Harbour Pond Mar 05, 2020 0.02 LTD LTD 3.30 Bartletts Harbour Bartletts Harbour Long Pond (same as Feb 26, 2020 0.02 LTD LTD 7.70 Castors River North) Bay L'Argent Bay L'Argent Sugarloaf Hill Pond Mar 03, 2020 0.01 LTD LTD 4.90 Bay Roberts Bay Roberts, Spaniard's Rocky Pond Feb 11, 2020 0.01 LTD LTD 3.00 Bay Bay de Verde Bay de Verde Island Pond Feb 18, 2020 0.01 LTD LTD 4.00 Beaches Beaches Grassey Pond Brook Mar 09, 2020 0.02 LTD LTD 7.70 Beachside Beachside Long Pond Mar 02, 2020 0.01 LTD LTD 6.00 Bellburns Bellburns Bound Brook Tributary Mar 10, 2020 0.07 0.19 LTD 4.50 Belleoram Belleoram Rabbits Pond Feb 04, 2020 0.01 LTD LTD 12.20 Bellevue Bellevue Big Pond Feb 26, 2020 0.03 LTD LTD 6.10 Bellevue Beach Bellevue Beach Unnamed Brook Feb 26, 2020 0.01 LTD LTD 6.80 Benton Benton