Bakalin V.A. 2016. a Revision of Lepidoziaceae (Hepaticae)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
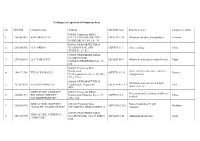
Catalogue of Exporters of Primorsky Krai № ITN/TIN Company Name Address OKVED Code Kind of Activity Country of Export 1 254308
Catalogue of exporters of Primorsky krai № ITN/TIN Company name Address OKVED Code Kind of activity Country of export 690002, Primorsky KRAI, 1 2543082433 KOR GROUP LLC CITY VLADIVOSTOK, PR-T OKVED:51.38 Wholesale of other food products Vietnam OSTRYAKOVA 5G, OF. 94 690001, PRIMORSKY KRAI, 2 2536266550 LLC "SEIKO" VLADIVOSTOK, STR. OKVED:51.7 Other ratailing China TUNGUS, 17, K.1 690003, PRIMORSKY KRAI, VLADIVOSTOK, 3 2531010610 LLC "FORTUNA" OKVED: 46.9 Wholesale trade in specialized stores China STREET UPPERPORTOVA, 38- 101 690003, Primorsky Krai, Vladivostok, Other activities auxiliary related to 4 2540172745 TEK ALVADIS LLC OKVED: 52.29 Panama Verkhneportovaya street, 38, office transportation 301 p-303 p 690088, PRIMORSKY KRAI, Wholesale trade of cars and light 5 2537074970 AVTOTRADING LLC Vladivostok, Zhigura, 46 OKVED: 45.11.1 USA motor vehicles 9KV JOINT-STOCK COMPANY 690091, Primorsky KRAI, Processing and preserving of fish and 6 2504001293 HOLDING COMPANY " Vladivostok, Pologaya Street, 53, OKVED:15.2 China seafood DALMOREPRODUKT " office 308 JOINT-STOCK COMPANY 692760, Primorsky Krai, Non-scheduled air freight 7 2502018358 OKVED:62.20.2 Moldova "AVIALIFT VLADIVOSTOK" CITYARTEM, MKR-N ORBIT, 4 transport 690039, PRIMORSKY KRAI JOINT-STOCK COMPANY 8 2543127290 VLADIVOSTOK, 16A-19 KIROV OKVED:27.42 Aluminum production Japan "ANKUVER" STR. 692760, EDGE OF PRIMORSKY Activities of catering establishments KRAI, for other types of catering JOINT-STOCK COMPANY CITYARTEM, STR. VLADIMIR 9 2502040579 "AEROMAR-ДВ" SAIBEL, 41 OKVED:56.29 China Production of bread and pastry, cakes 690014, Primorsky Krai, and pastries short-term storage JOINT-STOCK COMPANY VLADIVOSTOK, STR. PEOPLE 10 2504001550 "VLADHLEB" AVENUE 29 OKVED:10.71 China JOINT-STOCK COMPANY " MINING- METALLURGICAL 692446, PRIMORSKY KRAI COMPLEX DALNEGORSK AVENUE 50 Mining and processing of lead-zinc 11 2505008358 " DALPOLIMETALL " SUMMER OCTOBER 93 OKVED:07.29.5 ore Republic of Korea 692183, PRIMORSKY KRAI KRAI, KRASNOARMEYSKIY DISTRICT, JOINT-STOCK COMPANY " P. -

The Rediscovery and Conservation Status of Bazzania Bhutanica In
BryophytesAbroad Bazzania bhutanica in Bhutan The rediscovery and conservation status of Bazzania bhutanica in Bhutan Prior to 2009, only hutan is a small kingdom in the east n Tongsa Dzong, one of the splendid religious fortresses Himalayan Mountains sandwiched of Bhutan. D. Long one colony of Bazzania between India and China (Tibet), v B. bhutanica at one of its two known localities at Buduni near Samtse in south-west Bhutan, where it is bhutanica was known still relatively unexplored for critically endangered due to pressures of deforestation bryophytes, but nevertheless known from Bhutan. However, and development. D. Long Bto have a rich bryophyte flora. Mizutani (1967) during an investigation recorded six species of Bazzania from Bhutan, & Grolle, 1990), which increased the number although three of these [B. tridens (Reinw. et al.) of Bazzania species known from Bhutan to 14. last winter, a second Trevis., B. himalayana (Mitt.) Schiffn. and B. Study of the mosses has progressed more slowly, colony was discovered. sikkimensis (Steph.) Herz. were erroneous records but it is hoped to publish an updated moss originating from the Kalimpong district, West checklist soon. Many new records for Bhutan David G. Long, Bengal (former ‘British Bhutan’)]. His reports and a few species new to science emerged from Baboo Ram Gurung of B. imbricata (Mitt.) S.Hatt., B. griffithiana the liverwort collections; one of the latter was (Steph.) Mizutani and B. appendiculata (Mitt.) Bazzania bhutanica, discovered in 1982 and and Rebecca Pradhan S.Hatt. were based on specimens collected by published as a new species by Kitagawa & Grolle describe the story William Griffith in Bhutan in 1838. -

Lepidozia Bragginsiana, a New Species from New Zealand (Marchantiopsida)
Phytotaxa 173 (2): 117–126 ISSN 1179-3155 (print edition) www.mapress.com/phytotaxa/ PHYTOTAXA Copyright © 2014 Magnolia Press Article ISSN 1179-3163 (online edition) http://dx.doi.org/10.11646/phytotaxa.173.2.2 Lepidozia bragginsiana, a new species from New Zealand (Marchantiopsida) ENDYMION D. COOPER1 & MATT A.M. RENNER2 1Cell Biology and Molecular Genetics, 2107 Bioscience Research building, University of Maryland, College Park, MD 20742-4451, USA 2Royal Botanic Gardens & Domain Trust, Mrs Macquaries Road, Sydney, NSW 2000, Australia (corresponding author: [email protected]) Abstract Molecular and morphological data support the recognition of a new Lepidozia species related to L. pendulina and also endemic to New Zealand, which we dedicate to Dr John Braggins. Lepidozia bragginsiana can be distinguished from closely related and other similar species by its bipinnate branching, the narrow underleaf lobes, typically uniseriate toward their tip on both primary and secondary shoots, the asymmetric underleaves on primary shoots that are usually narrower than the stem and also possess basal spines and spurs, the production of spurs and spines, or even accessory lobes, on the postical margin of primary and secondary shoot leaves; and by the relatively small leaf cells with evenly thickened walls. Lepidozia bragginsiana is an inhabitant of hyper-humid forest habitats where it occupies elevated microsites on the forest floor. A lectotype is proposed for L. obtusiloba. Introduction The Lepidoziaceae Limpricht in Cohn (1877: 310) is perhaps the most comprehensively treated family within Australasian liverworts, having been subject to intensive and ongoing study and revision (e.g. Schuster 1980, 2000; Schuster & Engel 1987, 1996 Engel & Glenny 2008, Engel & Merrill 2004, Engel & Schuster 2001, Cooper et al. -

° 2013 Annual Report ° Conservation Projects in the Russian Far East
° 2013 AnnuAl report ° ConservAtion projeCts in the russiAn FAr eAst Office 409, 2 Petra Velikogo Street Vladivostok, Russia 690091 Tel: +7 (423) 220-50-53 Fax: +7 (423) 220-50-48 E-mail: [email protected] Web-site: www.fundphoenix.org Annual report 2013 [PHOENIX FUND] Annual report 2013 BACKGrounD South of the Russian Far East represents the only area in the world where the Amur tigers and leopards still exist in the wild. The species are listed as Endangered by the IUCN and are on CITES Appendix I for protection status. Protected under the Russian and international laws and regulations, these rare predator populations are still threatened by poaching, habitat destruction, prey depletion and conflicts with people. For sixteen years the Phoenix Fund, Russian environmental NGO, has been conducting anti-poaching and habitat protection, environmental education and outreach, monitoring of industrial projects, paying compensations for livestock depredation in order to keep stable the Amur tiger and leopard populations. Below, we are glad to present our final report describing the project activities between January 1 and December 30, 2013. The activities described below are the result of joint efforts of many organizations, both Russian and international, and invaluable contribution of our supporters! [PHOENIX FUND] Annual report 2013 news in tiGer poliCy The year 2013 was declared by Vladimir Putin behind bars. On July 2, 2013, a new article 258.1 as the Year of Environment Protection in Russia. was introduced in the Russian Criminal Code We are glad to witness the ongoing steps by that envisages criminal responsibility for Russian Government to protect Amur tigers and poaching, keeping, acquisition, storage, leopards from extinction. -

A Study of Branching Patterns in Bazzania (Jungermanniales, Lepidoziaceae)
Polish Botanical Journal 58(2): 481–489, 2013 DOI: 10.2478/pbj-2013-0042 A STUDY OF BRANCHING PATTERNS IN BAZZANIA (JUNGERMANNIALES, LEPIDOZIACEAE) DAVI D MEAGHER Abstract. Branching patterns in the leafy liverwort genus Bazzania Gray were investigated in twenty-five Australasian species. Terminal branching was found to be neither consistently sinistrorse nor consistently dextrorse, and neither consistently homodromous nor consistently antidromous. Microphyllous ventral-intercalary branches arising from stems usually have ‘siblings’ arising from main branches. The production of leafy ventral-intercalary branches is not necessarily associated with the termination of stems or main branches. These results are contrary to previous ideas about branching in Bazzania. Key words: Bazzania, branching, Lepidoziaceae, liverwort David Meagher, School of Botany, The University of Melbourne, Victoria, Australia; e-mail: [email protected] INTRO D UCTION Branching patterns in leafy liverworts have been SPECI M ENS EXA M INE D studied intensively since the pioneering studies by Leitgeb (1871) and have been used to separate fami- Bazzania accreta (Lehm. & Lindenb.) Trevis. – HO- 312267 lies, genera and subgenera. There are two univer- Bazzania adnexa (Lehm. & Lindenb.) Trevis. var. adnexa sally recognised fundamental modes of branching – DAM-1526 (MELU) in leafy liverworts: (1) terminal branching, in which Bazzania amblyphylla Meagher – CBG-9519195 branches develop close to the growing tip of the stem Bazzania corbieri (Stephani) Meagher – DAM-551 from a cortical cell, and (2) intercalary branching, in (MELU) which branches arise from the stem medulla, so that Bazzania densa (Sande Lac.) Schiffn. – DAM-1112 the developing branch ruptures the cortex, leaving (MELU) behind a collar of cortical cells around the base of the Bazzania fasciculata (Stephani) Meagher – NSW-605694 branch (Schuster 1984). -

About the Book the Format Acknowledgments
About the Book For more than ten years I have been working on a book on bryophyte ecology and was joined by Heinjo During, who has been very helpful in critiquing multiple versions of the chapters. But as the book progressed, the field of bryophyte ecology progressed faster. No chapter ever seemed to stay finished, hence the decision to publish online. Furthermore, rather than being a textbook, it is evolving into an encyclopedia that would be at least three volumes. Having reached the age when I could retire whenever I wanted to, I no longer needed be so concerned with the publish or perish paradigm. In keeping with the sharing nature of bryologists, and the need to educate the non-bryologists about the nature and role of bryophytes in the ecosystem, it seemed my personal goals could best be accomplished by publishing online. This has several advantages for me. I can choose the format I want, I can include lots of color images, and I can post chapters or parts of chapters as I complete them and update later if I find it important. Throughout the book I have posed questions. I have even attempt to offer hypotheses for many of these. It is my hope that these questions and hypotheses will inspire students of all ages to attempt to answer these. Some are simple and could even be done by elementary school children. Others are suitable for undergraduate projects. And some will take lifelong work or a large team of researchers around the world. Have fun with them! The Format The decision to publish Bryophyte Ecology as an ebook occurred after I had a publisher, and I am sure I have not thought of all the complexities of publishing as I complete things, rather than in the order of the planned organization. -

North American H&A Names
A very tentative and preliminary list of North American liverworts and hornworts, doubtless containing errors and omissions, but forming a basis for updating the spreadsheet of recognized genera and numbers of species, November 2010. Liverworts Blasiales Blasiaceae Blasia L. Blasia pusilla L. Fossombroniales Calyculariaceae Calycularia Mitt. Calycularia crispula Mitt. Calycularia laxa Lindb. & Arnell Fossombroniaceae Fossombronia Raddi Fossombronia alaskana Steere & Inoue Fossombronia brasiliensis Steph. Fossombronia cristula Austin Fossombronia foveolata Lindb. Fossombronia hispidissima Steph. Fossombronia lamellata Steph. Fossombronia macounii Austin Fossombronia marshii J. R. Bray & Stotler Fossombronia pusilla (L.) Dumort. Fossombronia longiseta (Austin) Austin Note: Fossombronia longiseta was based on a mixture of material belonging to three different species of Fossombronia; Schuster (1992a p. 395) lectotypified F. longiseta with the specimen of Austin, Hepaticae Boreali-Americani 118 at H. An SEM of one spore from this specimen was previously published by Scott and Pike (1988 fig. 19) and it is clearly F. pusilla. It is not at all clear why Doyle and Stotler (2006) apply the name to F. hispidissima. Fossombronia texana Lindb. Fossombronia wondraczekii (Corda) Dumort. Fossombronia zygospora R.M. Schust. Petalophyllum Nees & Gottsche ex Lehm. Petalophyllum ralfsii (Wilson) Nees & Gottsche ex Lehm. Moerckiaceae Moerckia Gottsche Moerckia blyttii (Moerch) Brockm. Moerckia hibernica (Hook.) Gottsche Pallaviciniaceae Pallavicinia A. Gray, nom. cons. Pallavicinia lyellii (Hook.) Carruth. Pelliaceae Pellia Raddi, nom. cons. Pellia appalachiana R.M. Schust. (pro hybr.) Pellia endiviifolia (Dicks.) Dumort. Pellia endiviifolia (Dicks.) Dumort. ssp. alpicola R.M. Schust. Pellia endiviifolia (Dicks.) Dumort. ssp. endiviifolia Pellia epiphylla (L.) Corda Pellia megaspora R.M. Schust. Pellia neesiana (Gottsche) Limpr. Pellia neesiana (Gottsche) Limpr. -

Amur Tiger Conservation in Russia in 2017
Amur Tiger Conservation in Russia in 2017 Progress report by Phoenix Fund January 1 – June 30, 2017 SMART In February 2015, the simultaneous count of Amur tigers and Amur leopards showed that about 523-540 Amur tigers occur today in the Russian Far East (comparing to 430-500 individuals recorded during the previous count in 2005). Same upward tendency was registered with the global population of Amur leopards, which numbers grew from 30 to 60-70 species in a decade. Despite sustained conservation efforts over recent years and encouraging recent monitoring results, the big cats still remain at risk due to poaching, logging, forest fires, and prey depletion. Every year the wild populations of Amur tigers and Amur leopards officially lose up to ten individuals due to poaching, collisions with vehicles and other causes of death. According to official statistics and trusted sources, as many as 11 Amur tigers died from January through June 2017. The ongoing alarming mortality in these species requires powerful and innovative solutions that leverage and build on existing capacity if we are to be successful in halting the loss of invaluable endangered wildlife. In this regard, thanks to continuous support from the Kolmarden Fundraising Foundation Phoenix continued implementing its complex conservation programme with the following objectives: 1) to reduce poaching of Amur tigers and their prey species and improve protection of their habitat; 2) to improve law enforcement efforts within federal-level protected areas; 3) and to raise people’s awareness about the state of, and the threats to, the Amur tiger population and involve the public in nature conservation actions. -

Kurzia Makinoana (Steph.) Grolle
DRAFT, Version 1.1 Draft Management Recommendations for slender clawleaf Kurzia makinoana (Steph.) Grolle Version 1.1 November 4, 1996 TABLE OF CONTENTS EXECUTIVE SUMMARY .................................................... 2 I. Natural History ........................................................... 3 A. Taxonomic/Nomenclatural History ...................................... 3 B. Species Description .................................................. 3 1. Morphology .................................................. 3 2. Reproductive Biology ........................................... 4 3. Ecology .................................................... 4 C. Range, Known Sites ................................................. 4 D. Habitat Characteristics and Species Abundance ............................. 5 II. Current Species Situation ................................................... 5 A. Why Species is Listed under Survey and Manage Standards and Guidelines ........ 5 B. Major Habitat and Viability Considerations ................................ 6 C. Threats to the Species ................................................ 6 D. Distribution Relative to Land Allocations ................................. 6 III. Management Goals and Objectives ........................................... 7 A. Management Goals for the Taxon ....................................... 7 B. Specific Objectives .................................................. 7 IV. Habitat Management ..................................................... 7 A. Lessons from History -

Notes on Early Land Plants Today. 54. a Transfer in Lepidoziaceae (Marchantiophyta)
Phytotaxa 167 (2): 218–219 ISSN 1179-3155 (print edition) www.mapress.com/phytotaxa/ PHYTOTAXA Copyright © 2014 Magnolia Press Correspondence ISSN 1179-3163 (online edition) http://dx.doi.org/10.11646/phytotaxa.167.2.13 Notes on Early Land Plants Today. 54. A transfer in Lepidoziaceae (Marchantiophyta) ENDYMION D. COOPER1, LARS SÖDERSTRÖM2, ANDERS HAGBORG3 & MATT VON KONRAT3 1 CMNS-Cell Biology and Molecular Genetics, 2107 Bioscience Research Building, University of Maryland, College Park, MD 20742- 4451, USA; [email protected]. 2Department of Biology, Norwegian University of Science and Technology, N-7491 Trondheim, Norway; [email protected] 3Department of Science and Education, The Field Museum, 1400 South Lake Shore Drive, Chicago, IL 60605–2496, USA; hagborg@ pobox.com, [email protected] When Cooper et al. (2013) reorganized species among genera in Lepidoziaceae Limpricht (1876: 310), one taxon, Lepidozia leratii Stephani (1922: 333), was mistakenly combined under Neolepidozia Fulford & Taylor (1959: 81). The transfer was not based on any morphological or molecular evidence placing it in Neolepidozia. On the contrary, molecular phylogenetic studies (Cooper et al., 2011, 2012; Heslewood & Brown, 2007) all place Lepidozia leratii in close proximity of Tricholepidozia pulcherrima (Stephani 1909: 600) E.D.Cooper in Cooper et al. (2013: 60) the type of Tricholepidozia (Schuster 1963: 256) E.D.Cooper in Cooper et al. (2013: 58). This error is corrected here. Tricholepidozia leratii (Steph.) E.D.Cooper, comb nov. Basionym:—Lepidozia leratii Steph., Sp. Hepat. (Stephani) 6: 333 (Stephani 1922). Type:—New Caledonia, summit of Mt Mou, July 1909, Lerat (PC-0102363, lectotype by Hürlimann 1985 [http://coldb.mnhn.fr/catalognumber/mnhn/pc/pc0102363]). -

CONSERVATION ACTION PLAN for the RUSSIAN FAR EAST ECOREGION COMPLEX Part 1
CONSERVATION ACTION PLAN FOR THE RUSSIAN FAR EAST ECOREGION COMPLEX Part 1. Biodiversity and socio-economic assessment Editors: Yuri Darman, WWF Russia Far Eastern Branch Vladimir Karakin, WWF Russia Far Eastern Branch Andrew Martynenko, Far Eastern National University Laura Williams, Environmental Consultant Prepared with funding from the WWF-Netherlands Action Network Program Vladivostok, Khabarovsk, Blagoveshensk, Birobidzhan 2003 TABLE OF CONTENTS CONSERVATION ACTION PLAN. Part 1. 1. INTRODUCTION 4 1.1. The Russian Far East Ecoregion Complex 4 1.2. Purpose and Methods of the Biodiversity and Socio-Economic 6 Assessment 1.3. The Ecoregion-Based Approach in the Russian Far East 8 2. THE RUSSIAN FAR EAST ECOREGION COMPLEX: 11 A BRIEF BIOLOGICAL OVERVIEW 2.1. Landscape Diversity 12 2.2. Hydrological Network 15 2.3. Climate 17 2.4. Flora 19 2.5. Fauna 23 3. BIOLOGICAL CONSERVATION IN THE RUSSIAN FAR EAST 29 ECOREGION COMPLEX: FOCAL SPECIES AND PROCESSES 3.1. Focal Species 30 3.2. Species of Special Concern 47 3.3 .Focal Processes and Phenomena 55 4. DETERMINING PRIORITY AREAS FOR CONSERVATION 59 4.1. Natural Zoning of the RFE Ecoregion Complex 59 4.2. Methods of Territorial Biodiversity Analysis 62 4.3. Conclusions of Territorial Analysis 69 4.4. Landscape Integrity and Representation Analysis of Priority Areas 71 5. OVERVIEW OF CURRENT PRACTICES IN BIODIVERSITY CONSERVATION 77 5.1. Legislative Basis for Biodiversity Conservation in the RFE 77 5.2. The System of Protected Areas in the RFE 81 5.3. Conventions and Agreements Related to Biodiversity Conservation 88 in the RFE 6. SOCIO-ECONOMIC INFLUENCES 90 6.1. -

Securing a Future for Amur Leopards and Tigers in Russia
Securing a Future for Amur Leopards and Tigers in Russia – VI 2018 Final Report Phoenix Fund 1 Securing a Future for Amur Leopards and Tigers in Russia – VI • 2018 Final Report TABLE OF CONTENTS Background ................................................................................................................................................... 2 Project Summary ........................................................................................................................................... 3 Project Activities............................................................................................................................................ 4 SMART in five protected areas .................................................................................................................. 4 Annual workshop for educators ................................................................................................................ 8 Education in Khasan, Lazo, Terney and Vladivostok ................................................................................. 9 Tiger Day in Primorye .............................................................................................................................. 11 Art Contest .............................................................................................................................................. 13 Photo credits: PRNCO “Tiger “Centre”, Far Eastern Operational Customs Office, Land of the Leopard National Park, Alexander Ratnikov, and children's paintings