The Bacillus Anthracis Cell Envelope: Composition, Physiological Role, and Clinical Relevance
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio In
microorganisms Review The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio in the Treatment of Obesity and Inflammatory Bowel disease Spase Stojanov 1,2, Aleš Berlec 1,2 and Borut Štrukelj 1,2,* 1 Faculty of Pharmacy, University of Ljubljana, SI-1000 Ljubljana, Slovenia; [email protected] (S.S.); [email protected] (A.B.) 2 Department of Biotechnology, Jožef Stefan Institute, SI-1000 Ljubljana, Slovenia * Correspondence: borut.strukelj@ffa.uni-lj.si Received: 16 September 2020; Accepted: 31 October 2020; Published: 1 November 2020 Abstract: The two most important bacterial phyla in the gastrointestinal tract, Firmicutes and Bacteroidetes, have gained much attention in recent years. The Firmicutes/Bacteroidetes (F/B) ratio is widely accepted to have an important influence in maintaining normal intestinal homeostasis. Increased or decreased F/B ratio is regarded as dysbiosis, whereby the former is usually observed with obesity, and the latter with inflammatory bowel disease (IBD). Probiotics as live microorganisms can confer health benefits to the host when administered in adequate amounts. There is considerable evidence of their nutritional and immunosuppressive properties including reports that elucidate the association of probiotics with the F/B ratio, obesity, and IBD. Orally administered probiotics can contribute to the restoration of dysbiotic microbiota and to the prevention of obesity or IBD. However, as the effects of different probiotics on the F/B ratio differ, selecting the appropriate species or mixture is crucial. The most commonly tested probiotics for modifying the F/B ratio and treating obesity and IBD are from the genus Lactobacillus. In this paper, we review the effects of probiotics on the F/B ratio that lead to weight loss or immunosuppression. -

Amerithrax Investigative Summary
The United States Department of Justice AMERITHRAX INVESTIGATIVE SUMMARY Released Pursuant to the Freedom of Information Act Friday, February 19, 2010 TABLE OF CONTENTS I. THE ANTHRAX LETTER ATTACKS . .1 II. EXECUTIVE SUMMARY . 4 A. Overview of the Amerithrax Investigation . .4 B. The Elimination of Dr. Steven J. Hatfill as a Suspect . .6 C. Summary of the Investigation of Dr. Bruce E. Ivins . 6 D. Summary of Evidence from the Investigation Implicating Dr. Ivins . .8 III. THE AMERITHRAX INVESTIGATION . 11 A. Introduction . .11 B. The Investigation Prior to the Scientific Conclusions in 2007 . 12 1. Early investigation of the letters and envelopes . .12 2. Preliminary scientific testing of the Bacillus anthracis spore powder . .13 3. Early scientific findings and conclusions . .14 4. Continuing investigative efforts . 16 5. Assessing individual suspects . .17 6. Dr. Steven J. Hatfill . .19 7. Simultaneous investigative initiatives . .21 C. The Genetic Analysis . .23 IV. THE EVIDENCE AGAINST DR. BRUCE E. IVINS . 25 A. Introduction . .25 B. Background of Dr. Ivins . .25 C. Opportunity, Access and Ability . 26 1. The creation of RMR-1029 – Dr. Ivins’s flask . .26 2. RMR-1029 is the source of the murder weapon . 28 3. Dr. Ivins’s suspicious lab hours just before each mailing . .29 4. Others with access to RMR-1029 have been ruled out . .33 5. Dr. Ivins’s considerable skill and familiarity with the necessary equipment . 36 D. Motive . .38 1. Dr. Ivins’s life’s work appeared destined for failure, absent an unexpected event . .39 2. Dr. Ivins was being subjected to increasing public criticism for his work . -

Investigation of Anthrax Associated with Intentional Exposure
October 19, 2001 / Vol. 50 / No. 41 889 Update: Investigation of Anthrax Associated with Intentional Exposure and Interim Public Health Guidelines, October 2001 893 Recognition of Illness Associated with the Intentional Release of a Biologic Agent 897 Weekly Update: West Nile Virus Activity — United States, October 10–16, 2001 Update: Investigation of Anthrax Associated with Intentional Exposure and Interim Public Health Guidelines, October 2001 On October 4, 2001, CDC and state and local public health authorities reported a case of inhalational anthrax in Florida (1 ). Additional cases of anthrax subsequently have been reported from Florida and New York City. This report updates the findings of these case investigations, which indicate that infections were caused by the intentional release of Bacillus anthracis. This report also includes interim guidelines for postexposure pro- phylaxis for prevention of inhalational anthrax and other information to assist epidemi- ologists, clinicians, and laboratorians responding to intentional anthrax exposures. For these investigations, a confirmed case of anthrax was defined as 1) a clinically compatible case of cutaneous, inhalational, or gastrointestinal illness* that is laboratory confirmed by isolation of B. anthracis from an affected tissue or site or 2) other labora- tory evidence of B. anthracis infection based on at least two supportive laboratory tests. A suspected case was defined as 1) a clinically compatible case of illness without isola- tion of B. anthracis and no alternative diagnosis, but with laboratory evidence of B. anthracis by one supportive laboratory test or 2) a clinically compatible case of an- thrax epidemiologically linked to a confirmed environmental exposure, but without cor- roborative laboratory evidence of B. -

2001 Anthrax Letters
Chapter Five: 2001 Anthrax Letters Author’s Note: The analysis and comments regarding the communication efforts described in this case study are solely those of the authors; this analysis does not represent the official position of the FDA. This case was selected because it is one of the few major federal efforts to distribute medical countermeasures in response to an acute biological incident. These events occurred more than a decade ago and represent the early stages of US biosecurity preparedness and response; however, this incident serves as an excellent illustration of the types of communication challenges expected in these scenarios. Due in part to the extended time since these events and the limited accessibility of individual communications and messages, this case study does not provide a comprehensive assessment of all communication efforts. In contrast to the previous case studies in this casebook, the FDA’s role in the 2001 anthrax response was relatively small, and as such, this analysis focuses principally on the communication efforts of the CDC and state and local public health agencies. The 2001 anthrax attacks have been studied extensively, and the myriad of internal and external assessments led to numerous changes to response and communications policies and protocols. The authors intend to use this case study as a means of highlighting communication challenges strictly within the context of this incident, not to evaluate the success or merit of changes made as a result of these events. Abstract The dissemination of Bacillus anthracis via the US Postal Service (USPS) in 2001 represented a new public health threat, the first intentional exposure to anthrax in the United States. -

Distribution and Characteristics of Bacillus Bacteria Associated with Hydrobionts and the Waters of the Peter the Great Bay, Sea of Japan I
ISSN 0026-2617, Microbiology, 2008, Vol. 77, No. 4, pp. 497–503. © Pleiades Publishing, Ltd., 2008. Original Russian Text © I.A. Beleneva, 2008, published in Mikrobiologiya, 2008, Vol. 77, No. 4, pp. 558–565. EXPERIMENTAL ARTICLES Distribution and Characteristics of Bacillus Bacteria Associated with Hydrobionts and the Waters of the Peter the Great Bay, Sea of Japan I. A. Beleneva1 Zhirmunskii Institute of Marine Biology, Far East Division, Russian Academy of Sciences, ul. Pal’chevskogo, 17, Vladivostok 690041, Russia Received May 28, 2007 Abstract—Bacilli of the species Bacillus subtilis, B. pumilus, B. mycoides, B. marinus and B. licheniformis (a total of 53 strains) were isolated from 15 invertebrate species and the water of the Vostok Bay, Peter the Great Bay, Sea of Japan. Bacilli were most often isolated from bivalves (22.7%) and sea cucumbers (18.9%); they occurred less frequently in sea urchins and starfish (13.2 and 7.5%, respectively). Most of bacilli strains were isolated from invertebrates inhabiting silted sediments. No Bacillus spp. strains were isolated from invertebrates inhabiting stony and sandy environments. The species diversity of bacilli isolated from marine objects under study was low. Almost all bacterial isolates were resistant to lincomycin. Unlike B. pumilus, B. subtilis isolates were mostly resistant to benzylpenicillin and ampicillin. Antibiotic sensitivity of B. licheniformis strains was variable (two strains were resistant to benzylpenicillin and oxacillin, while one was sensitive). A significant fraction of isolated bacilli contained pigments. Pigmented strains were more often isolated from seawater sam- ples, while colorless ones predominated within hydrobionts. B. subtilis colonies had the broadest range of co- lors. -

Bacillus Cereus Pneumonia
‘Advances in Medicine and Biology’, Volume 67, 2013 Nova Science Publisher Inc., Editor: Berhardt LV To Alessia, my queen, and Giorgia, my princess Chapter BACILLUS CEREUS PNEUMONIA Vincenzo Savini* Clinical Microbiology and Virology, Spirito Santo Hospital, Pescara (PE), Italy ABSTRACT Bacillus cereus is a Gram positive/Gram variable environmental rod, that is emerging as a respiratory pathogen; particularly, pneumonia it causes may be serious and can resemble the anthrax disease. In fact, the organism is strictly related to the famous Bacillus anthracis, with which it shares genotypical, fenotypical, and pathogenic features. Treatment of B. cereus lower airway infections is becoming increasingly hard, due to the spread of multidrug resistance traits among members of the species. Hence, the present chapter’s scope is to shed a light on this bacterium’s lung pathogenicity, by depicting salient microbiological, epidemiological and clinical features it shows. Also, we would like to explore virulence determinants and resistance mechanisms that make B. cereus a life-threatening, potentially difficult-to-treat agent of airway pathologies. * Email: [email protected]. 2 Vincenzo Savini INTRODUCTION The genus Bacillus includes spore-forming species, strains of which are not usually considered to be clinically relevant when isolated from human specimens; in fact, these bacteria are known to be ubiquitously distributed in the environment and may easily contaminate culture material along with improperly handled sample collection devices (Miller, 2012; Brooks, 2001). Bacillus anthracis is the prominent agent of human pathologies within the genus Bacillus, although it is uncommon in most clinical laboratories (Miller, 2012). It is a frank pathogen, as it causes skin and enteric infections; above all, however, it is responsible for a serious lung disease (the ‘anthrax’) that is frequently associated with bloodstream infection and a high mortality rate (Frankard, 2004). -

Lessons from the Anthrax Attacks Implications for US
Lessons from the Anthrax Attacks Implications for US. Bioterrorism Preparedness A Report on a National Forum on Biodefense Author David Heymart Research Assistants Srusha Ac h terb erg, L Joelle Laszld Organized by the Center for Strategic and International Studies and the Defense Threat Reduction Agency --CFr”V? --.....a DlSTRf BUTION This report ISfor official use only; distribution authorized to U S. government agencies, designated contractors, and those with an official need Contains information that may be exempt from public release under the Freedom of Information Act. exemption number 2 (5 USC 552); exemption number 3 (’lo USC 130).Approvalof the Defense Threat Reduction Agency prior to public release is requrred Contract Number OTRAM-02-C-0013 For Official Use Only About CSIS For four decades, the Center for Stravgic and Internahonal Studies (CSIS)has been dedicated 10 providing world leaders with strategic insights on-and pohcy solubons tcurrentand emergtng global lssues CSIS IS led by John J Hamre, former L S deputy secretary of defense It is guided by a board of trustees chaired by former U S senator Sam Nunn and consistlng of prominent individuals horn both the public and private sectors The CSIS staff of 190 researchers and support staff focus pnrnardy on three subjecr areas First, CSIS addresses the fuU spectrum of new challenges to national and mternabonal security Second, it maintains resident experts on all of the world's myor geographical regions Third, it IS committed to helping to develop new methods of governance for the global age, to this end, CSIS has programs on technology and pubhc policy, International trade and finance, and energy Headquartered in Washington, D-C ,CSIS IS pnvate, bipartlsan, and tax-exempt CSIS docs not take specific policy positions, accordygly, all views expressed herein should be understood to be solely those ofthe author Spousor. -

Chapter 11: Bacteria Bacterial Groups
Bacterial Groups u Most widely accepted taxonomic classification for bacteria is Bergey’s Manual of Systematic Bacteriology. u 5000 bacterial species identified, 3100 classified. Chapter 11: Bacteria u Bacteria are divided into four divisions (phyla) according to the characteristics of their cell walls. u Each division is divided into sections according to: u Gram stain reaction u Cell shape u Cell arrangements u Oxygen requirements u Motility u Nutritional and metabolic properties u Each section contains several genera. Four Divisions of Bacteria Classification of Bacteria Procaryotes Gram-Negative Division II Wall-Less Archaea Bacteria Bacteria Bacteria Bacteria (Gracilicutes) (Firmicutes) (Tenericutes) (Mendosicutes) Thin Cell Walls Thick cell Walls Lack cell walls Unusual cell walls Division I. Gram-Negative Bacteria Gram Negative Bacteria Spirochetes 1. Spirochetes u Helical shape. Flexible. u Contain two or more axial filaments (endoflagella). u Move in corkscrew pattern. u Medically important members: F Treponema pallidum: Syphilis F Borrelia spp.: Lyme disease, relapsing fever F Leptospira: Leptospirosis 1 Syphilis is Caused by a Spirochete Lyme Disease is Caused by a Spirochete Primary syphilitic chancre and secondary rash. Source: Tropical Medicine and Parasitology, 1997 Lyme Disease early lesion at tick bite site. Source: Medical Microbiology, 1998 2. Aerobic, Motile, Helical/Vibroid Gram- Negative Bacteria Gram Negative Bacteria u Rigid helical shape or curved rods. Aerobic, Motile, Helical/Vibroid u Lack axial filaments (endoflagella); have polar Gram-Negative Bacteria flagella instead. u Most are harmless aquatic organisms. u Genus Azospirillum fixes nitrogen in soil. u Genus Bdellovibrio attacks other bacteria. u Important pathogens include: F Campylobacter jejuni: Most common bacterial food- borne intestinal disease in the United States (2 million cases/year). -

Technical Method
J Clin Pathol: first published as 10.1136/jcp.38.9.1073 on 1 September 1985. Downloaded from J Clin Pathol 1985;38:1073-1084 Technical method Use of microwaves for acid and alcohol fast staining S HAFIZ, RC SPENCER, MARGARET LEE, organism depends on several factors but remains an HILARY GOOCH, BI DUERDEN Department important component in the identification of a ofMedical Microbiology, Sheffield University Medi- restricted group of organisms. cal School, Sheffield The technical problems associated with the classic Ziehl-Neelsen method include the dangers of heat- ing by flaming torch in laboratories in which volatile Certain bacteria that are characterised by a high organic solvents are used, the deposits that accumu- content of lipid in their cell walls cannot be stained late on the underside of the slide as a result of heat- by simple stains, and either heat or prolonged con- ing, and the crystalline deposits of stain that collect tact is required to drive the stain into the cells; once on the film as a result of evaporation and drying. stained they resist decolourisation by mineral acids This method also requires 20-30 minutes of or acid and alcohol. These organisms are designated laboratory time. Numerous minor modifications acid fast or acid and alcohol fast. They include have been made to the Ziehl-Neelsen method by Mycobacterium tuberculosis and related mycobac- increasing the concentration of carbolic magenta or teria, Mycobacterium leprae, and certain of the by cold staining techniques,'0-'2 but the basic prob- actinomycetes. Koch first stained the tubercle bacil- lems remain. lus by immersion in an alkaline solution of Some of the problems may be overcome by the copyright. -
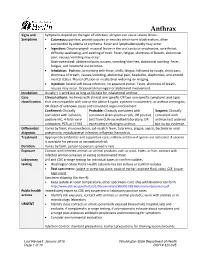
Anthrax Reporting and Investigation Guideline
Anthrax Signs and Symptoms depend on the type of infection; all types can cause severe illness: Symptoms • Cutaneous: painless, pruritic papules or vesicles which form black eschars, often surrounded by edema or erythema. Fever and lymphadenopathy may occur. • Ingestion: Oropharyngeal: mucosal lesion in the oral cavity or oropharynx, sore throat, difficulty swallowing, and swelling of neck. Fever, fatigue, shortness of breath, abdominal pain, nausea/vomiting may occur. Gastrointestinal: abdominal pain, nausea, vomiting/diarrhea, abdominal swelling. Fever, fatigue, and headache are common. • Inhalation: Biphasic, presenting with fever, chills, fatigue, followed by cough, chest pain, shortness of breath, nausea/vomiting, abdominal pain, headache, diaphoresis, and altered mental status. Pleural effusion or mediastinal widening on imaging. • Injection: Severe soft tissue infection; no apparent eschar. Fever, shortness of breath, nausea may occur. Occasional meningeal or abdominal involvement. Incubation Usually < 1 week but as long as 60 days for inhalational anthrax Case Clinical criteria: An illness with at least one specific OR two non-specific symptoms and signs classification that are compatible with one of the above 4 types, systemic involvement, or anthrax meningitis; OR death of unknown cause and consistent organ involvement Confirmed: Clinically Probable: Clinically consistent with Suspect: Clinically consistent with isolation, consistent Gram-positive rods, OR positive consistent with positive IHC, 4-fold rise in test from CLIA-accredited laboratory, OR anthrax test ordered antibodies, PCR, or LF MS epi evidence relating to anthrax but no epi evidence Differential Varies by form; mononucleosis, cat-scratch fever, tularemia, plague, sepsis, bacterial or viral diagnosis pneumonia, mycobacterial infection, influenza, hantavirus Treatment Appropriate antibiotics and supportive care; anthrax antitoxin if spores are activated. -

Bioterrorism, Biological Weapons and Anthrax
Bioterrorism, Biological Weapons and Anthrax Part IV Written by Arthur H. Garrison Criminal Justice Planning Coordinator Delaware Criminal Justice Council Bioterrorism and biological weapons The use of bio-terrorism and bio-warfare dates back to 6th century when the Assyrians poisoned the well water of their enemies. The goal of using biological weapons is to cause massive sickness or death in the intended target. Bioterrorism and biological weapons The U.S. took the threat of biological weapons attack seriously after Gulf War. Anthrax vaccinations of U.S. troops Investigating Iraq and its biological weapons capacity The Soviet Union manufactured various types of biological weapons during the 1980’s • To be used after a nuclear exchange • Manufacturing new biological weapons – Gene engineering – creating new types of viruses/bacteria • Contagious viruses – Ebola, Marburg (Filoviruses) - Hemorrhagic fever diseases (vascular system dissolves) – Smallpox The spread of biological weapons after the fall of the Soviet Union •Material • Knowledge and expertise •Equipment Bioterrorism and biological weapons There are two basic categories of biological warfare agents. Microorganisms • living organic germs, such as anthrax (bacillus anthrax). –Bacteria –Viruses Toxins • By-products of living organisms (natural poisons) such as botulism (botulinum toxin) which is a by- product of growing the microorganism clostridium botulinum Bioterrorism and biological weapons The U.S. was a leader in the early research on biological weapons Research on making -

Antibacterial Photosensitization Through Activation of PNAS PLUS Coproporphyrinogen Oxidase
Antibacterial photosensitization through activation of PNAS PLUS coproporphyrinogen oxidase Matthew C. Surdela, Dennis J. Horvath Jr.a, Lisa J. Lojeka, Audra R. Fullena, Jocelyn Simpsona, Brendan F. Dutterb,c, Kenneth J. Sallenga, Jeremy B. Fordd, J. Logan Jenkinsd, Raju Nagarajane, Pedro L. Teixeiraf, Matthew Albertollec,g, Ivelin S. Georgieva,e,h, E. Duco Jansend, Gary A. Sulikowskib,c, D. Borden Lacya,d, Harry A. Daileyi,j,k, and Eric P. Skaara,1 aDepartment of Pathology, Microbiology, and Immunology, Vanderbilt University Medical Center, Nashville, TN 37232; bDepartment of Chemistry, Vanderbilt University, Nashville, TN 37232; cVanderbilt Institute for Chemical Biology, Nashville, TN 37232; dDepartment of Biomedical Engineering, Vanderbilt University, Nashville, TN 37232; eVanderbilt Vaccine Center, Vanderbilt University Medical Center, Nashville, TN 37232; fBiomedical Informatics, Vanderbilt University School of Medicine, Nashville, TN 37203; gDepartment of Biochemistry, Vanderbilt University, Nashville, TN 37232; hDepartment of Electrical Engineering and Computer Science, Vanderbilt University, Nashville, TN 37232; iBiomedical and Health Sciences Institute, University of Georgia, Athens, GA 30602; jDepartment of Microbiology, University of Georgia, Athens, GA 30602; and kDepartment of Biochemistry and Molecular Biology, University of Georgia, Athens, GA 30602 Edited by Ferric C. Fang, University of Washington School of Medicine, Seattle, WA, and accepted by Editorial Board Member Carl F. Nathan June 26, 2017 (received for review January 10, 2017) Gram-positive bacteria cause the majority of skin and soft tissue Small-molecule VU0038882 (‘882) was previously identified in infections (SSTIs), resulting in the most common reason for clinic a screen for activators of the S. aureus heme-sensing system two- visits in the United States.