Smooth Newt, Lissotriton Vulgaris Ampelensis Fuhn 1951, in Romania
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Bid-Book Brasov 2021 - European Capital of Culture Brasov 2021 Team
MUNICIPALITY OF BRASOV Bid-Book Brasov 2021 - European Capital of Culture Brasov 2021 Team: Project coordinator: DRAGOS DAVID Concept, vision & artistic strategy: NICOLAE PEPENE The team: NICOLAE PEPENE RADU COLȚ SORIN GANEA GABRIELA BRAȘOVEANU MEDEEA KATERINA PETROVAN Design & editing: INOVATIV MEDIA Translations: Eran Catt Photo credits: Agenția Metropolitană Brașov Primăria Municipiului Brașov Revista ASTRA Nicolae Pepene Asociația Carpaterra / Mihail Iacomir Biblioteca Județeană G. Barițiu Brașov Emi Cristea Victor Ștefănescu Inovativ Media Municipality of Brasov Bid-Book Brasov 2021 - European Capital of Culture In Brasov County, not afar from the old medieval road between Brasov and Sighisoara, at Mercheasa (in the commune named Homorod), 60 km from the city of Brasov, stands ”The Elder of the Carpathians“, a mountain oak (sessile), whose age is twice that of the two prominent historic towns of Romania’s past. Brasov’s historic coat of arms is a royal crown placed on the cut trunk of amountain oak (sessile) extended in 13 strong roots. Ever since the Middle Ages, throughout Europe, the oak symbolizes solidity, strength, longevity and height, both in the spiritual and in the material sense. It has always and everywhere represented both moral and physical strength. Also, through its crown, the oak is a symbol of hospitality and it represented the equivalent of a temple in the European art. It is the oak that the people of Brasov have chosen in 2015 as a symbol to define the past, present and near future of the city they inhabit. 3 Introduction – General considerations Brasov has chosen to grow through culture! The city located on the border lined by the The first documentary attestation Carpathian Mountains on the border of the kingdom, of Brasov happened in 1235, empire or principality, protected its riches behind strong fortifications, erected upon the urge of King Sigismund under the name of Corona. -

Strategy for Local Sustainable Development of the City of Codlea 2011-2020
Innovation in administration EUROPEAN UNION GOVERNMENT OF ROMANIA European Social Fund Operational programme Ministry of Administration and "Administrative Capacity Interior Development" STRATEGY FOR LOCAL SUSTAINABLE DEVELOPMENT OF THE CITY OF CODLEA 2011-2020 -SUMMARY- Territorial and Administrative Unit “City of Codlea” 1 During 15 months the City of Codlea carried out the project called “Update of strategy for local sustainable development of the locality of Codlea”. The project was co-financed from the EUROPEAN SOCIAL FUND through the Operational programme for Administrative Capacity Development 2007-2013, project SMIS code 12802. By implementing the project, several results were obtained, the most important being the document “Strategy for local sustainable development of the municipality of Codlea”, as approved by the Codlea Local Council. Other results: 4 specialty studies (economic, social, technical–economic and environmental studies) which ensured the grounding of municipality’s development strategy; SWOT analysis; Codlea Town Hall staff trained in the fields of strategic planning, sustainable development and equal opportunities; Specific software for strategy management and implementation within the territorial and administrative unit of the municipality of Codlea – results monitoring; People informed about the content of strategy – document published on the institution’s website. Part II STRATEGY OF DEVELOPMENT Chapter I. Vision of development The main approach within the update process of strategy for sustainable development of the City of Codlea was the intercommunity one. The city of Codlea is a part of the Brasov Metropolitan Area, alongside the cities of Brasov, Săcele, Râşnov, Predeal, Ghimbav, the localities of Sânpetru, Bod, Hălchiu, Prejmer, Tărlungeni, Cristian, Vulcan, Hărman, Feldioara and Crizbav. -
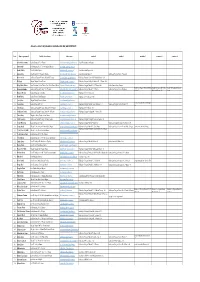
2019 2020 Consilieri Scolari
Adrese de contact ale profesorilor consilieri scolari din judetul BRASOV Nr.crt. Numele si prenumele Unitatea de invatamant Adresa email arondare 1 arondare 2 arondare 3 arondare 4 arondare 5 1 Alexandrescu Cristina Școala Gimnazială Nr. 14 Brașov [email protected] Școala Gimnazială nr. 16 Brașov 2 Andraș Alina Școala Gimnazială Nr. 9 'Nicolae Orghidan' Brașov [email protected] 3 Barbu Mihaela Liceul 'Șt.O.Iosif' Rupea [email protected] Școala Gimnazială Rupea - 588 4 Bârsan Crina Liceul Teoretic 'I.C. Drăgușanu' Victoria [email protected] Școala Gimnazială Voila 115 Grădinița cu Program Normal Voivodeni 5 Bica Cornelia Grădinița cu Program Prelungit 'Prichindeii' Făgăraș [email protected] Grădinița cu Program Săptămânal 'Pinochio' Făgăraș - 281 6 Blaj Ligia Colegiul Național 'Unirea' Brașov [email protected] Grădinița cu Program Prelungit 'Martinică' Nr. 17 Brașov- 164 7 Blejan Maria Liliana Liceul Vocațional de Arte Plastice 'Hans Mattis-Teutsch' Brașov [email protected] Grădinița cu Program Normal Nr. 17C Brașov-200 Școala Gimnazială nr. 4 Brașov Grădinița cu Program Normal Sâmbăta Școala Gimnazială 'Ion Codru Școala Gimnazială Sâmbăta de 8 Bordeanu Luminița Grădinița cu Program Prelungit Nr. 1 Victoria [email protected] Grădinița cu Program Normal Nr. 2 Victoria Grădinița cu Program Normal Drăguș-48 de Sus-47 Drăgușanu' Drăguș- 48 Sus-81 9 Botezatu Mariana Școala Gimnazială Nr. 2 Codlea [email protected] Gradinita cu PP nr. 1 Codlea - 233 10 Bratu Raluca Scoala Gimaziala Vama Buzaului [email protected] Gradinita cu PP nr. 2 Brasov - 300 11 Carțiș Silvia Colegiul 'Nicolae Titulescu' Brașov [email protected] Școala Gimnazială Lunca Câlnicului - 12 Casian Alina Școala Gimnazială Prejmer [email protected] Grădinița cu Program Normal Lunca Câlnicului- 113 Grădinița cu Program Normal Prejmer- 69 256 13 Cîdă Simona Grădinița cu Program Prelungit 'Boboceii' Nr. -

Razing of Romania's Past.Pdf
REPORT Ttf F1 *t 'A. Í M A onp DlNU C GlURESCU THE RAZING OF ROMANIA'S PAST The Razing of Romania's Past was sponsored by the Kress Foundation European Preservation Program of the World Monuments Fund; it was published by USACOMOS. The World Monuments Fund is a U.S. nonprofit organization based in New York City whose purpose is to preserve the cultural heritage of mankind through administration of field restora tion programs, technical studies, advocacy and public education worldwide. World Monuments Fund, 174 East 80th Street, New York, N.Y. 10021. (212) 517-9367. The Samuel H. Kress Foundation is a U.S. private foundation based in New York City which concentrates its resources on the support of education and training in art history, advanced training in conservation and historic preservation in Western Europe. The Samuel H. Kress Foundation, 174 East 80th Street, N.Y. 10021. (212) 861-4993. The United States Committee of the International Council on Monuments and Sites (USACOMOS) is one of 60 national committees of ICOMOS forming a worldwide alliance for the study and conservation of historic buildings, districts and sites. It is an international, nongovernmental institution which serves as the focus of international cultural resources ex change in the United States. US/ICOMOS, 1600 H Street, N.W., Washington, D.C., 20006. (202) 842-1866. The text and materials assembled by Dinu C. Giurescu reflect the views of the author as sup ported by his independent research. Book design by DR Pollard and Associates, Inc. Jacket design by John T. Engeman. Printed by J.D. -

Scenarii Unitati De Invatamant
Nr.crt. Unitatea școlară Tip UAT Localitatea Statut Scenariul 1 Școala Gimnazială 'Apaczai Csere Janos' Apața de stat Apața Apața PJ 1 2 Grădinița cu Program Normal Apața de stat Apața Apața structură 1 3 Școala Gimnazială Augustin de stat Augustin Augustin PJ 1 4 Grădinița cu Program Normal Augustin de stat Augustin Augustin structură 1 5 Școala Primară Beclean de stat Beclean Beclean PJ 1 6 Grădinița cu Program Normal Beclean de stat Beclean Beclean structură 1 7 Grădinița cu Program Normal Boholț de stat Beclean Boholț structură 1 8 Școala Primară Boholț de stat Beclean Boholț structură 1 9Școala Gimnazială Bod de stat Bod Bod PJ 1 10 Grădinița cu Program Normal Bod de stat Bod Bod structură 1 11Liceul 'Sextil Pușcariu' Bran de stat Bran Bran PJ 2 12 Grădinița cu Program Normal Bran de stat Bran Bran structură 2 13 Grădinița cu Program Normal Poarta de stat Bran Poarta structură 2 14Școala Primară Poarta de stat Bran Poarta structură 2 15 Grădinița cu Program Normal Predeluț de stat Bran Predeluț structură 2 16 Grădinița cu Program Normal Sohodol de stat Bran Sohodol structură 2 17 Școala Gimnazială Sohodol de stat Bran Sohodol structură 2 18 Grădinița cu Program Normal Șimon de stat Bran Șimon structură 2 19 Școala Gimnazială Șimon de stat Bran Șimon structură 2 20 Centrul Școlar pentru Educație Incluzivă Brașov de stat Brașov Brașov PJ 2 21 Centrul Școlar pentru Educație Incluzivă de Recuperare Hipoacuzici Brașov de stat Brașov Brașov structură 2 22 Grădinița Specială Brașov de stat Brașov Brașov structură 2 23 Clubul Sportiv Brașovia Brașov de stat Brașov Brașov PJ 2 24 Clubul Sportiv Dinamo Brașov de stat Brașov Brașov PJ 2 25 Colegiul de Științe ale Naturii 'Emil Racoviță' Brașov de stat Brașov Brașov PJ 2 26 Colegiul de Științe 'Grigore Antipa' Brașov de stat Brașov Brașov PJ 2 27 Colegiul Național 'Andrei Șaguna' Brașov de stat Brașov Brașov PJ 2 28 Colegiul Național 'Aprily Lajos' Brașov de stat Brașov Brașov PJ 2 29 Grădinița cu Program Normal 'Aprily Lajos' Brașov de stat Brașov Brașov structură 2 30 Colegiul Național de Informatică 'Gr. -

Brasov Could Achieve Energy Savings in the ‘Public Transport’ Sector (By Modernizing the Bus Fleet, Purchasing Energy Efficient Rolling Stock, Etc.)
1 The findings, interpretations, and conclusions expressed in this report do not necessarily reflect the views and position of the Table of Contents Executive Directors of the World Bank, the European Union, or the Government of Romania. Executive Summary /1 The TRACE diagnostic is part of work done under the Romania Regional Development Program – a Reimbursable Advisory Background /5 Service activity, undertaken by the World Bank at the request National Legislation Regarding Energy /5 of the Ministry of Regional Development and Public Background Brașov /11 Administration, with EU funding. The report was written by a team comprised of Manuela Mot, Ranjan Bose, Sebastian Sustainable Brașov /16 Burduja, and Marcel Ionescu-Heroiu. Cristina Zirimis has Street Lighting/16 provided logistical and administrative support throughout the Municipal Buildings /18 process. The team would like to thank the colleagues at the Power /20 Ministry of Regional Development and Public Administration Urban Transport/22 (particularly Ionuţ Trincă and Bogdan Ţigău), as well as the District Heating/29 colleagues in the Brașov City Hall and the Brașov County Water and Waste Water/33 Council, who supported the team throughout. Solid Waste/36 TRACE (Tool for Rapid Assessment of City Energy) was developed by ESMAP (Energy Sector Management Assistance Energy Efficiency Recommendations /40 Program), a unit of the World Bank, and is available for District Heating /41 download and free use at: http://esmap.org/TRACE Urban Transport /44 Municipal Buildings /51 Street Lighting /53 Water/55 Annexes /57 1 Executive Summary in the transport sector include: parking, modernization of the bus fleet, After the 1989 Revolution, Romania began its transition from a centralized and the development of non-motorized transport by building more bike system to a market based economy. -

Nume Localitate Strada Nr Denumirea Actului
Nume Localitate Strada Nr Denumirea actului INSTIINTARI DEPLATA PF 2017 AANICAI GHIORGHE BRASOV DACIA 69 Instiintare de plata 2017 ABABEI DENIS‐TITUS ROMAN APRODUL ARBORE Instiintare de plata 2017 ABDEL‐RAZEQ MAHMUD BRASOV 15 NOIEMBRIE80 Instiintare de plata 2017 ACASANDREI VASILE LUNCA CALNICUI.C FRIMU 419 Instiintare de plata 2017 ACATRINEI ADRIAN PREJMER UZINEI 836FInstiintare de plata 2017 ACATRINEI ELENA‐ALINA PREJMER UZINEI 836FInstiintare de plata 2017 ACETER MIRUNA SILVIA BRASOV ALBATROSULU13 Instiintare de plata 2017 ACUCULITEI ADRIAN BRASOV CERNATULUI 14A Instiintare de plata 2017 ADAM ALEXANDRU LUNCA CALNICUI.C FRIMU 425 Instiintare de plata 2017 ADAM CONSTANTIN MARIAN PREJMER MORII 993 Instiintare de plata 2017 ADAM GEORGETA LUNCA MARCUSULUI 15 Instiintare de plata 2017 ADAM IOAN LUNCA CALNICUPRINCIPALA 75 B Instiintare de plata 2017 ADAM IOAN LUNCA CALNICUMICA 59 Instiintare de plata 2017 ADAM IONUT ALEXANDRU LUNCA CALNICUPRINCIPALA 79B Instiintare de plata 2017 ADAM MARIA LUNCA CALNICUI.C FRIMU 425 Instiintare de plata 2017 ADAM MARIA LUNCA MARCUSBISERICII 61 Instiintare de plata 2017 AGA IULIAN CHICHIS/BACEL Instiintare de plata 2017 AGA STEFAN MINDRA 179 Instiintare de plata 2017 AGYO ANISOARA LUNCA CALNICUI.C FRIMU 417 Instiintare de plata 2017 AGYO FERENC LUNCA CALNICUI.C FRIMU 417 Instiintare de plata 2017 AILENEI CONSTANTIN LUNCA CALNICUPRINCIPALA 55 Instiintare de plata 2017 AILENEI CONSTANTIN MUGURELUNCA CALNICUPRINCIPALA 55 Instiintare de plata 2017 AILENEI IOAN‐RAZVAN LUNCA CALNICUPRINCIPALA 55 Instiintare de plata -

Report on Territorial Context Analysis
Report on Territorial context analysis Partner n. 7 BRASOV COUNTY ... a trail brought me here Doc. 6A-TCA-P7 28.12.2018 Mod . 5B-3T-5 1 Table of contents Table of contents ................................................................................................................................... 2 1. General Description of Brașov County Region and its Demography............................................. 3 2. Economy and Labour Market ........................................................................................................ 7 3. Environment and Climate Change ............................................................................................... 10 4. Infrastructure and Mobility ......................................................................................................... 13 5. Tourism and Leisure .................................................................................................................... 18 6. Existing policies related to ThreeT project features. Projects .................................................... 29 7. Stakeholders ................................................................................................................................ 31 8. S.W.O.T. Analysis ......................................................................................................................... 35 Annexes 1-2-3-4 Mod . 5B-3T-5 2 1. General Description of Brașov County Region and its Demography Situated in the central part of Romania, in the south-eastern part of Transylvania, -

Locuri Vacante Pentru Etapa a Ii-A
Inspectoratul Școlar Județean Brașov Admiterea în clasa a IX-a LOCURI VACANTE PENTRU ETAPA A II-A Liceu ZI Cod Unitate Localitate Profil Specializare Limba Bilingv Locuri 114 Colegiul Național 'Aprily Lajos' Brașov Brașov Real Matematică-Informatică maghiară 2 113 Colegiul Național 'Aprily Lajos' Brașov Brașov Umanist Filologie maghiară Engleză 15 112 Colegiul Național 'Aprily Lajos' Brașov Brașov Servicii Comerţ maghiară 12 136 Colegiul Național de Informatică 'Gr. Moisil' Brașov Brașov Real Matematică-Informatică 1 105 Colegiul 'Nicolae Titulescu' Brașov Brașov Real Ştiinţe ale Naturii 1 107 Colegiul 'Nicolae Titulescu' Brașov Brașov Umanist Ştiinţe Sociale 1 103 Colegiul 'Nicolae Titulescu' Brașov Brașov Umanist Filologie 1 157 Colegiul Tehnic de Transporturi Brașov Brașov Tehnic Electronică automatizări 20 158 Colegiul Tehnic de Transporturi Brașov Brașov Tehnic Producţie media 8 156 Colegiul Tehnic Energetic 'Remus Răduleț' Brașov Brașov Tehnic Electronică automatizări 18 153 Colegiul Tehnic Energetic 'Remus Răduleț' Brașov Brașov Tehnic Electric 18 138 Colegiul Tehnic 'Maria Baiulescu' Brașov Brașov Servicii Estetica şi igiena corpului omenesc 1 142 Colegiul Tehnic 'Mircea Cristea' Brașov Brașov Tehnic Construcţii, inst. şi lucrări publice 7 152 Colegiul Tehnic 'Transilvania' Brașov Brașov Tehnic Mecanică 25 186 Liceul 'Andrei Mureșanu' Brașov (Seminarul Teologic) Brașov Teologic Teologie ortodoxă 6 -- Liceul cu Program Sportiv Brașov Brașov Sportiv Sportiv 4 -- Liceul Vocațional de Muzică 'Tudor Ciortea' Brașov Brașov Artistic Muzică 3 185 Liceul Tehnologic Silvic 'Dr. Nicolae Rucăreanu' Brașov Brașov Resurse Silvicultură 9 148 Colegiul Tehnic 'Simion Mehedinți' Codlea Codlea Servicii Economic 5 150 Colegiul Tehnic 'Simion Mehedinți' Codlea Codlea Servicii Turism şi alimentaţie 14 118 Colegiul Național 'Doamna Stanca' Făgăraș Făgăraș Real Ştiinţe ale Naturii 1 117 Colegiul Național 'Doamna Stanca' Făgăraș Făgăraș Real Ştiinţe ale Naturii germană 13 177 Liceul Tehnologic 'Dr. -

Designing and Implementing an Urban Mobility Plan in Brasov, Romania …A Brief Analysis
Designing and implementing an urban mobility plan in Brasov, Romania …a brief analysis Mr. Catalin Frangulea-Pastor, Programme Director, Brasov Metropolitan Agency for Sustainable Development • Context • The process of SUMP elaboration • Challenges in the implementation of the SUMP CONTEXT The Brasov Growth pole includes 3 municipalities (Brasov, Codlea, Sacele), 4 cities (Ghimbav, Predeal, Rasnov and Zarnesti) and 11 rural communes (Bod, Budila, Cristian, Crizbav, Feldioara, Halchiu, Hă rman, Prejmer, Sânpetru, Tarlungeni and Vulcan) The Growth pole has a total population of 400.000 inhabitants and a surface area of 139 628 ha What is a SUMP? A Sustainable Urban Mobility Plan seeks to contribute to development of an urban transport systemwhich: (a.) Is accessible and meets the basic mobility needs of all users; (b.) Balances and responds to the diverse demands for mobility and transport services by citizens, businesses and industry; (c.) Guides a balanced development and better integration of the different transport modes; (d.) Meets the requirements of sustainability, balancing the need for economic viability, social equity, health and environmentalquality; (e.) Optimises efficiency and cost effectiveness; (f.) Makes better use of urban space and of existing transport infrastructure and services; (g.) Enhances the attractiveness of the urban environment, quality of life, and public health; (h.) Improves traffic safety and security; (i.) Reduces air and noise pollution, greenhouse gas emissions, and energy consumption; (j.) Contributes to a better overall performance of the trans-European transport network and the Europe's transport systemas a whole. SUMP elaboration $$$$ Brasov SUMP – an overview The consortium of international consultants was let by WSP PARSONS- BRINKERHOFF The traffic data has been collected in December 2014 – February 2015 The implementation process started in late 2016 and is currently under way. -

Research Findings on Social Capital and Matters in Brasov Community
Bulletin of the Transilvania University of Braşov Series VII: Social Sciences • Law • Vol. 6 (55) No. 2 - 2013 RESEARCH FINDINGS ON SOCIAL CAPITAL AND MATTERS IN BRASOV COMMUNITY. A SECONDARY ANALYSIS APPROACH Claudiu COMAN1 Abstract: The aim of this article is to present new research findings regarding sustainable development projects in Brasov County, depending on the social matters and social capital. The method used is a secondary analysis of data collected in a 2009 survey carried out on 3960 persons, representative for Brasov County. This new approach manages to reveal a classification of the Brasov County localities according to the social capital typology (bonding or bridging) and according to the identified social matters’ categories. The data indicates new perspectives on the two types of social capital and on their relation to social matters, representing guidelines for future research. Key words: social capital, social matters, secondary analysis, bonding social capital, bridging social capital. 1. Introduction: conceptual with their values and measures must be clarifications taken to the end of eliminating such a situation [18]. These authors underline the Social matters are not rooted in the following defining elements of social social circumstances but in the matters: presumptions of the interested parties and 1. The situation presumed to be of the partisans committed to win over the problematic. The existence of the political acceptance in the sense of situations is supported by people talking perceiving certain circumstances as about it, by mass-media airing it; problems, depending on which said 2. The inconsistency of such a situation perceptions reflects the evolution of the with certain values and interests. -

ITEMA 2020Conference Proceedings
THE MARKETING MIX - A BASIC TOOL IN THE MANAGEMENT OF THE TOURIST DESTINATION Delia Fratu1 DOI: https://doi.org/10.31410/ITEMA.2020.55 Abstract: Kotler's 2002 definition of place marketing is as follows: "Place marketing means designing a site to meet the needs of its target markets. The development of places means the development for a place of a systematic long-term marketing strategy, directed towards the identification and development of the natural and potential attributes of an area”. The elaboration of a marketing mix for the tourist destination is the main tool for marketing planning and for adopting the most appropriate strategies. Given that each tourist destination has its specificity, the marketing mix must contain its own elements, adapted to the objectives set. In this paper we chose to exemplify a way to apply and combine the components of the following marketing mix: product, price, distribution, promotion, staff, partnerships, packaging, security, process management for the tourist destination Brasov Area. Keywords: Place marketing, Marketing mix, Tourism, Destination. 1. BRASOV AREA- A TOURIST DESTINATION tourist destination is defined according to the following variables: the attractions and the environment within the destination, the facilities and services available at the A destination, the accessibility of the destination, the image and the brand of the destination. 1.1. Defining the Tourist Destination Brasov Area Brasov Area is located in a special natural setting. In the southern part it is delimited by the Făgăraş, Bucegi, Piatra Mare, Piatra Craiului and Postăvaru Mountains, in the Eastern part there is the Braşov depression, and in the Western part it is bordered by the Olt river valley.