Bacterial Lipoteichoic Acid Attenuates Toll-Like Receptor Dependent Dendritic Cells Activation and Inflammatory Response
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
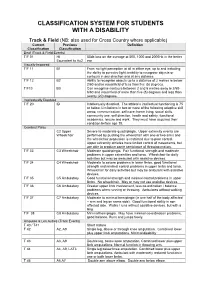
Disability Classification System
CLASSIFICATION SYSTEM FOR STUDENTS WITH A DISABILITY Track & Field (NB: also used for Cross Country where applicable) Current Previous Definition Classification Classification Deaf (Track & Field Events) T/F 01 HI 55db loss on the average at 500, 1000 and 2000Hz in the better Equivalent to Au2 ear Visually Impaired T/F 11 B1 From no light perception at all in either eye, up to and including the ability to perceive light; inability to recognise objects or contours in any direction and at any distance. T/F 12 B2 Ability to recognise objects up to a distance of 2 metres ie below 2/60 and/or visual field of less than five (5) degrees. T/F13 B3 Can recognise contours between 2 and 6 metres away ie 2/60- 6/60 and visual field of more than five (5) degrees and less than twenty (20) degrees. Intellectually Disabled T/F 20 ID Intellectually disabled. The athlete’s intellectual functioning is 75 or below. Limitations in two or more of the following adaptive skill areas; communication, self-care; home living, social skills, community use, self direction, health and safety, functional academics, leisure and work. They must have acquired their condition before age 18. Cerebral Palsy C2 Upper Severe to moderate quadriplegia. Upper extremity events are Wheelchair performed by pushing the wheelchair with one or two arms and the wheelchair propulsion is restricted due to poor control. Upper extremity athletes have limited control of movements, but are able to produce some semblance of throwing motion. T/F 33 C3 Wheelchair Moderate quadriplegia. Fair functional strength and moderate problems in upper extremities and torso. -

Fish-Stream Identification Guidebook
of BRITISH COLUMBIA Fish-stream Identification Guidebook Second edition Version 2.1 August 1998 BC Environment Fish-stream Identification Guidebook of BRITISH COLUMBIA Fish-stream Identification Guidebook Second edition Version 2.1 August 1998 Authority Forest Practices Code of British Columbia Act Operational Planning Regulation Canadian Cataloguing in Publication Data Main entry under title: Fish-stream identification guidebook. – 2nd ed. (Forest practices code of British Columbia) ISBN 0-7726-3664-8 1. Fishes – Habitat – British Columbia. 2. River surveys – British Columbia. 3. Forest management – British Columbia. 4. Riparian forests – British Columbia – Management. I. British Columbia. Ministry of Forests. SH177.L63F58 1998 634.9 C98-960250-8 Fish-stream Identification Guidebook Preface This guidebook has been prepared to help forest resource managers plan, prescribe and implement sound forest practices that comply with the Forest Practices Code. Guidebooks are one of the four components of the Forest Practices Code. The others are the Forest Practices Code of British Columbia Act, the regulations, and the standards. The Forest Practices Code of British Columbia Act is the legislative umbrella authorizing the Code’s other components. It enables the Code, establishes mandatory requirements for planning and forest practices, sets enforcement and penalty provisions, and specifies administrative arrangements. The regulations lay out the forest practices that apply province-wide. The chief forester may establish standards, where required, to expand on a regulation. Both regulations and standards are mandatory requirements under the Code. Forest Practices Code guidebooks have been developed to support the regulations, however, only those portions of guidebooks cited in regulation are part of the legislation. -

1 ICAM3-Fc Outperforms Receptor-Specific Antibodies
Preprints (www.preprints.org) | NOT PEER-REVIEWED | Posted: 10 April 2019 doi:10.20944/preprints201904.0118.v1 Peer-reviewed version available at Molecules 2019, 24, 1825; doi:10.3390/molecules24091825 ICAM3-Fc outperforms receptor-specific antibodies targeted nanoparticles to dendritic cells for cross-presentation Luis J. Cruz,1* Paul J. Tacken,2 Johan S. van der Schoot,2 Felix Rueda,3 Ruurd Torensma,2 Carl G. Figdor2* 1Translational Nanobiomaterials and Imaging, Department of Radiology, Leiden University Medical Center, Albinusdreef 2, 2333 ZA Leiden, The Netherlands. 2Department of Tumor Immunology, Radboud Insititute for Molecular Life Sciences, Radboud University Medical Center, Postbox 9101, 6500 HB Nijmegen, The Netherlands. 3Department of Biochemistry and Molecular Biology, University of Barcelona, Diagonal 643, 08028 Barcelona, Spain. Running title: ICAM3-Fc- versus antibody-targeted NP vaccines to human DCs Keywords: Delivery system; nanoparticle; targeting. AUTHOR INFORMATION Corresponding Author *Dr. Luis J. Cruz Translational Nanobiomaterials and Imaging, Department of Radiology, Bldg.1, C5-60. Leiden University Medical Center, Albinusdreef 2 2333 ZA Leiden, The Netherlands Tel: +31 71 5263025 Email: [email protected] *Prof. Dr. Carl G. Figdor Department of Tumor Immunology, Nijmegen Centre for Molecular Life Sciences, Radboud University Nijmegen Medical Centre, Postbox 9101, 6500 HB Nijmegen, The Netherlands. Fax: +31-24-3540339; Tel.: +31-24-3617600; E-mail: [email protected] 1 © 2019 by the author(s). Distributed -

Synoptic-Scale Control Over Modern Rainfall and Flood Patterns in the Levant Drylands with Implications for Past Climates
JUNE 2018 ARMONETAL. 1077 Synoptic-Scale Control over Modern Rainfall and Flood Patterns in the Levant Drylands with Implications for Past Climates MOSHE ARMON Fredy and Nadine Herrmann Institute of Earth Sciences, Hebrew University of Jerusalem, Givat Ram, Jerusalem, Israel ELAD DENTE Fredy and Nadine Herrmann Institute of Earth Sciences, Hebrew University of Jerusalem, Givat Ram, and Geological Survey of Israel, Jerusalem, Israel JAMES A. SMITH Department of Civil and Environmental Engineering, Princeton University, Princeton, New Jersey YEHOUDA ENZEL AND EFRAT MORIN Fredy and Nadine Herrmann Institute of Earth Sciences, Hebrew University of Jerusalem, Givat Ram, Jerusalem, Israel (Manuscript received 23 January 2018, in final form 1 May 2018) ABSTRACT Rainfall in the Levant drylands is scarce but can potentially generate high-magnitude flash floods. Rainstorms are caused by distinct synoptic-scale circulation patterns: Mediterranean cyclone (MC), active Red Sea trough (ARST), and subtropical jet stream (STJ) disturbances, also termed tropical plumes (TPs). The unique spatiotemporal char- acteristics of rainstorms and floods for each circulation pattern were identified. Meteorological reanalyses, quantitative precipitation estimates from weather radars, hydrological data, and indicators of geomorphic changes from remote sensing imagery were used to characterize the chain of hydrometeorological processes leading to distinct flood patterns in the region. Significant differences in the hydrometeorology of these three flood-producing synoptic systems were identified: MC storms draw moisture from the Mediterranean and generate moderate rainfall in the northern part of the region. ARST and TP storms transfer large amounts of moisture from the south, which is converted to rainfall in the hyperarid southernmost parts of the Levant. -

The Use of Next Generation Sequencing to Study the Environmental Mycobiome and Its Potential Health Effects
The use of next generation sequencing to study the environmental mycobiome and its potential health effects Emma Marczylo Bioaerosols – June 2017 Overview • Brief Background: • Why are CRCE interested in fungal bioaerosols? • Mycobiome analysis: • Why use next generation sequencing? • Ongoing work • What is the current focus of CRCE’s bioaerosol research? 2 Bioaerosols – June 2017 Why bioaerosols? • Respiratory health/toxicology a big focus within our department & bioaerosols represent a current respiratory health concern 3 Bioaerosols – June 2017 Why bioaerosols? • Common sources relevant to public health 4 Bioaerosols – June 2017 Why bioaerosols? • Public concern over health effects of living near composting and intensive farming sites • Systematic reviews on exposures and health outcomes related to bioaerosol emissions from composting facilities (published 2015*) or intensive farming (ongoing) in collaboration with SAHSU • Evidence on both exposure assessment and health effects are limited • A big unknown is the microbial composition of such bioaerosols *Pearson et al, 2015, J Toxicol Environ Health B Crit Rev,18:43-69 5 Bioaerosols – June 2017 Why fungi? Normally die rapidly due to water evaporation, although increased humidity and clumping can prolong survival • Can remain viable for much longer periods, even at low humidity & high/low temperatures • Much less known about the fungal composition of bioaerosols (and other samples) • Fungi linked with development and exacerbation of asthma symptoms 6 Bioaerosols – June 2017 Mycobiome -

Framework for In-Field Analyses of Performance and Sub-Technique Selection in Standing Para Cross-Country Skiers
sensors Article Framework for In-Field Analyses of Performance and Sub-Technique Selection in Standing Para Cross-Country Skiers Camilla H. Carlsen 1,*, Julia Kathrin Baumgart 1, Jan Kocbach 1,2, Pål Haugnes 1 , Evy M. B. Paulussen 1,3 and Øyvind Sandbakk 1 1 Centre for Elite Sports Research, Department of Neuromedicine and Movement Science, Faculty of Medicine and Health Sciences, Norwegian University of Science and Technology, 7491 Trondheim, Norway; [email protected] (J.K.B.); [email protected] (J.K.); [email protected] (P.H.); [email protected] (E.M.B.P.); [email protected] (Ø.S.) 2 NORCE Norwegian Research Centre AS, 5008 Bergen, Norway 3 Faculty of Health, Medicine & Life Sciences, Maastricht University, 6200 MD Maastricht, The Netherlands * Correspondence: [email protected]; Tel.: +47-452-40-788 Abstract: Our aims were to evaluate the feasibility of a framework based on micro-sensor technology for in-field analyses of performance and sub-technique selection in Para cross-country (XC) skiing by using it to compare these parameters between elite standing Para (two men; one woman) and able- bodied (AB) (three men; four women) XC skiers during a classical skiing race. The data from a global navigation satellite system and inertial measurement unit were integrated to compare time loss and selected sub-techniques as a function of speed. Compared to male/female AB skiers, male/female Para skiers displayed 19/14% slower average speed with the largest time loss (65 ± 36/35 ± 6 s/lap) Citation: Carlsen, C.H.; Kathrin found in uphill terrain. -

Field Indicators of Hydric Soils
United States Department of Field Indicators of Agriculture Natural Resources Hydric Soils in the Conservation Service United States In cooperation with A Guide for Identifying and Delineating the National Technical Committee for Hydric Soils Hydric Soils, Version 8.2, 2018 Field Indicators of Hydric Soils in the United States A Guide for Identifying and Delineating Hydric Soils Version 8.2, 2018 (Including revisions to versions 8.0 and 8.1) United States Department of Agriculture, Natural Resources Conservation Service, in cooperation with the National Technical Committee for Hydric Soils Edited by L.M. Vasilas, Soil Scientist, NRCS, Washington, DC; G.W. Hurt, Soil Scientist, University of Florida, Gainesville, FL; and J.F. Berkowitz, Soil Scientist, USACE, Vicksburg, MS ii In accordance with Federal civil rights law and U.S. Department of Agriculture (USDA) civil rights regulations and policies, the USDA, its Agencies, offices, and employees, and institutions participating in or administering USDA programs are prohibited from discriminating based on race, color, national origin, religion, sex, gender identity (including gender expression), sexual orientation, disability, age, marital status, family/parental status, income derived from a public assistance program, political beliefs, or reprisal or retaliation for prior civil rights activity, in any program or activity conducted or funded by USDA (not all bases apply to all programs). Remedies and complaint filing deadlines vary by program or incident. Persons with disabilities who require alternative means of communication for program information (e.g., Braille, large print, audiotape, American Sign Language, etc.) should contact the responsible Agency or USDA’s TARGET Center at (202) 720-2600 (voice and TTY) or contact USDA through the Federal Relay Service at (800) 877-8339. -

Class II MHC Cytoplasmic Domain-Mediated Signaling in B Cells a Tail of Two Signals
Human Immunology 80 (2019) 32–36 Contents lists available at ScienceDirect Human Immunology journal homepage: www.elsevier.com/locate/humimm Class II MHC cytoplasmic domain-mediated signaling in B cells: A tail of two T signals Jonathan A. Harton Department of Immunology & Microbial Disease, Albany Medical College, 47 New Scotland Avenue, MC-151, Albany, NY 12208, USA ARTICLE INFO ABSTRACT Keywords: In addition to their role in antigen presentation, class II MHC molecules also transmit signals to B lymphocytes. MHC class II Class II MHC-mediated signals initiate a range of events in B cells, including induction of cell surface proteins, HLA-D initiation of cell-cycle progression/proliferation, activation of or protection from apoptosis, and antigen-de- Signaling pendent plasma cell differentiation. Although various transmembrane signaling proteins associate with classII B cells MHC molecules, the class II MHC cytoplasmic domains are essential for signals leading to increased intracellular cAMP and activation of protein kinase C (PKC). Although truncation and mutagenesis studies have provided considerable information about the cytoplasmic domain sequences required, how class II MHC molecules elicit cAMP and PKC activation is not known. Further, appropriate T-dependent B cell responses require intact cAMP and PKC signaling, but the extent to which class II MHC signals are involved is also unknown. This review details our current knowledge of class II MHC cytoplasmic domain signaling in B cells with an emphasis on the likely importance of class II MHC signals for T-dependent antibody responses. 1. Introduction complex, is only found on CD4+ T cells which are dependent upon thymic MHC class II for their development. -

Cell Biology from an Immune Perspective in This Lecture We Will
Harvard-MIT Division of Health Sciences and Technology HST.176: Cellular and Molecular Immunology Course Director: Dr. Shiv Pillai Cell Biology from an Immune Perspective In this lecture we will very briefly review some aspects of cell biology which are required as background knowledge in order to understand how the immune system works. These will include: 1. A brief overview of protein trafficking 2. Signal transduction 3. The cell cycle Some of these issues will be treated in greater depth in later lectures. Protein Trafficking/The Secretory Pathway: From an immune perspective the secretory compartment and structures enclosed by vesicles are “seen” in different ways from proteins that reside in the cytosol or the nucleus. We will briefly review the secretory and endocytic pathways and discuss the biogenesis of membrane proteins. Some of the issues that will be discussed are summarized in Figures 1-3. Early endosomes Late endosomes Lysosomes Golgi Vesiculo-Tubular Compartment ER Figure 1. An overview of the secretory pathway Early endosomes Late endosomes Multivesicular and multilamellar bodies Golgi ER Proteasomes Figure 2. Protein degradation occurs mainly in lysosomes and proteasomes Proteins that enter the cell from the environment are primarily degraded in lysosomes. Most cytosolic and nuclear proteins are degraded in organelles called proteasomes. Intriguingly these two sites of degradation are each functionally linked to distinct antigen presentation pathways, different kinds of MHC molecules and the activation of different categories of T cells. Integral membrane proteins maybe inserted into the membrane in a number of ways, the two most common of these ways being considered in Figure 3. -

To the New City of Brockville Zoning By-Law
BROCKVILLE FREQUENTLY ASKED QUESTIONS | SUMMARY OF CHANGES | HOW TO USE THE NEW ZONING BY-LAW GUIDE TO THE NEW CITY OF BROCKVILLE ZONING BY-LAW FEBRUARY 2014 TABLE OF CONTENTS FREQUENTLY ASKED What is a Zoning By-law? . 2 QUESTIONS Why is the City updating its Zoning By-law? . 2 What is the City of Brockville’s Official Plan? . 2 What does the City’s Official Plan say about the P .2 Zoning By-law? . 2 How does the City enforce and implement its Zoning By-law? 3 How will the Zoning By-law update affect me? . 4 HOW HAS THE What if my zoning did change? Will the buildings and the use of my property continue to be legal? . 4 ZONING BY-LAW Have you recently applied for or received approval for a CHANGED? planning application or building permit? . 4 Modernized, expanded definitions . 7 P .5 Improved format and organization . 8 Special provisions and design requirements for the Downtown and Central Waterfront Area – New Schedule “B” . 9 HOW DO I New zones for future neighbourhoods . 10 USE THE NEW Other changes to the Zoning Map (Schedule “A”) . 11 ZONING BY-LAW? Structure of the Zoning By-law . 12 How to check your zoning and identify P .12 Applicable Regulations . 13 HOW DO I How do I provide input on the Draft Zoning By-law? . 18 PROVIDE INPUT ON THE DRAFT ZONING BY-LAW P .18 FREQUENTLY ASKED QUESTIONS WHY IS THE CITY UPDATING ITS ZONING BY-LAW? The City is updating its Zoning By-law because the City’s new Official Plan has been WHAT IS A ZONING approved and is now in-effect . -

MHC Class II Stabilization at the Surface of Human Dendritic Cells Is the Result of Maturation-Dependent MARCH I Down-Regulation
MHC class II stabilization at the surface of human dendritic cells is the result of maturation-dependent MARCH I down-regulation Aude De Gassart*†‡, Voahirana Camosseto*†‡, Jacques Thibodeau§¶, Maurizio Ceppi*†‡, Nadia Catalan*†‡, Philippe Pierre*†‡ʈ, and Evelina Gatti*†‡ʈ *Centre d’Immunologie de Marseille–Luminy, Universite´ delaMe´ diterrane´e, Case 906, 13288 Marseille Cedex 9, France; †Institut National de la Sante´etde la Recherche Me´dicale, Unite´631, 13288 Marseille, France; ‡Centre National de la Recherche Scientifique, Unite´Mixte de Recherche 6102, 13288 Marseille, France; §De´partement de Microbiologie et Immunologie, Universite´de Montre´al, Montreal, QC, Canada H3T 1J4; and ¶Institut National de la Sante´et de la Recherche Me´dicale, Unite´743, Montreal, QC, Canada H3T 1J4 Edited by Ralph M. Steinman, The Rockefeller University, New York, NY, and approved January 9, 2008 (received for review September 19, 2007) In response to Toll-like receptor ligands, dendritic cells (DCs) dramatically when cells are exposed to LPS (12, 13). As determined dramatically enhance their antigen presentation capacity by sta- by confocal microscopy, iDCs accumulate HLA-DR molecules in bilizing at the cell-surface MHC II molecules. We demonstrate here LAMP1ϩ late endosomes and lysosomes. On activation by LPS, that, in human monocyte-derived DCs, the RING-CH ubiquitin E3 rapid relocalization of MHC II to the cell surface is observed (4–8 ligase, membrane-associated RING-CH I (MARCH I), promotes the h) [supporting information (SI) Fig. 7A]. Recently, this process has ubiquitination of the HLA-DR -chain. Thus, in nonactivated DCs, been shown to be associated with a loss of ubiquitination on the MARCH I induces the surface internalization of mature HLA-DR class II -chain in mouse bone marrow-derived DCs (bmDCs) (4, complexes, therefore reducing their stability and levels. -

ELIGIBILITY Para-Cycling Athletes: Must Be a United States Citizen With
ELIGIBILITY Para-cycling Athletes: Must be a United States citizen with a USA racing nationality. LICENSING National Championships: Riders may have a current International or Domestic USA Cycling license (USA citizenship) or Foreign Federation license showing a USA racing nationality to register. World Championships Selection: Riders must have a current International USA Cycling license with a USA racing nationality on or before June 20, 2019 in order to be selected for the Team USA roster for the 2019 UCI Para-cycling Road World Championships. Selection procedures for the World Championships can be found on the U.S. Paralympics Cycling Website: https://www.teamusa.org/US- Paralympics/Sports/Cycling/Selection-Procedures REGULATIONS General: All events conducted under UCI Regulations, including UCI equipment regulations. Road Race and Time Trials: • No National Team Kit or National championship uniforms are allowed. • For the Road Race, only neutral service and official’s cars are allowed in the caravan. • For the Time Trial, bicycles and handcycles must be checked 15 minutes before the athlete’s assigned start time. Courtesy checks will be available from 1 hour before the first start. No follow vehicles are allowed. • For all sport classes in the road race, athletes are required to wear a helmet in the correct sport class color, or use an appropriately color helmet cover, as follows: RED MC5, WC5, MT2, MH4, WH4, MB WHITE MC4, WC4, MH3, WH3, WB, WT2 BLUE MC3, WC3, MH2, WT1 BLACK MH5, WH5, MC2, WC2, MT1 YELLOW MC1, WC1, WH2 GREEN MH1 ORANGE WH1 Handcycle Team Relay (TR): New National Championship event run under UCI and special regulations below: • Team Requirements: Teams eligible for the National Championship Team Relay, must be respect the following composition: o Teams of three athletes o Using the table below, the total of points for the three TR athletes may not be more than six (6) points which must include an athlete with a scoring point value of 1.