Thesis Chapter 8
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

01 2019-07-23 Planning and Regulatory Functions Minutes.Pdf
ITEM 1 North Yorkshire County Council Planning and Regulatory Functions Committee Minutes of the meeting held at County Hall, Northallerton on 23 July 2019 at 10.00 am. Present:- County Councillors Peter Sowray (Chairman), David Blades, Eric Broadbent, Robert Heseltine, David Hugill, Mike Jordan, John McCartney, Richard Musgrave, Chris Pearson, and Clive Pearson. County Councillor Stuart Martin was in attendance. There were 3 members of the public present. Copies of all documents considered are in the Minute Book 89. Minutes Resolved - That the Minutes of the meeting held on 2 April 2019, having been printed and circulated, be taken as read and confirmed and signed by the Chairman as a correct record. 90. Declarations of Interest There were no declarations of interest. 91. Public Questions or Statements The representative of the Assistant Chief Executive (Legal and Democratic Services) stated that, apart from the people who had registered to speak in respect of the applications below, and who would be invited to do so during consideration of those Items, there were no questions or statements from members of the public. 92. C4/19/00523/CMA – (NY/2019/0016/73A) - Variation of Condition Nos. 19 & 20 & of Planning Permission Ref: C6/500/109/F/CMA to enable Asphalt Operations to continue until 21:00 hours Monday to Friday until 31 December 2021 at Pateley Bridge Quarry (Coldstones), Greenhow Hill, Pateley Bridge, Harrogate Considered - The report of the Corporate Director - Business and Environmental Services requesting Members to determine a planning application to enable asphalt operations and HGV movements to continue until 21:00 hours Monday to Friday (from the current end time of 18:00 hours) until 31 December 2021 at Pateley Bridge Quarry. -

Abramson, P, Berg DS, Fossick MR Roman Castleford
Abramson, P, Berg D S, Fossick M R Roman Castleford: Volume 2, The Structural and Environmental Evidence Castleford 17 2000 35 (Book Review ) Abramson, P A Penny for Your Forts Yorkshire 5 Winter 1987-8 20 Abramson, P Late Iron Age Settlement at Scotch Corner Scotch Corner, North Yorkshire 13 1996 8 Adamson, C H Burn's Cottage, Greta Bridge: An archaeological watching brief Greta Bridge, Durham 15 1998 6 Allon, P A future for the Roman Fort at Ilkley? Ilkley 4 Winter 1986-7 18 Annis, R Two Romano-British Settlement sites at Coulby Newham, Cleveland Coulby Newham, Cleveland 10 1992-3 9 Archaeological Services (WYAS) Recent Excavations by Archaeological Services (WYAS) 14 1997 20 Archaeological Services (WYAS) Excavations in 1997 by Archaeological Services (WYAS) 15 1998 13 Archaeological Services (WYAS) Excavations in 1998 by Archaeological Services (WYAS) 16 1999 18 Archaeological Services (WYAS) Moss Carr Wood, Methley, Leeds Methley 19 2002-2003 20 Archaeological Services (WYAS) Ferrybridge Ferrybridge 19 2002-2003 21 Archaeological Services (WYAS) Ilkley Ilkley 19 2002-2003 21 Archaeological Services (WYAS) North Leeds North Leeds 19 2002-2003 21 Archaeological Services (WYAS) Barnburgh Hall, Barnburgh, South Yorkshire Barnburgh 19 2002-2003 21 Archaeological Services (WYAS) West Moor Park Armthorpe, Doncaster Armthorpe, Doncaster 19 2002-2003 21 Archaeological Services (WYAS) High Street, Shafton, South Yorkshire Shafton 19 2002-2003 21 Archaeological Services (WYAS) Topham Farm, Sykehouse, South Yorkshire Sykehouse 19 2002-2003 22 -

CEF - Covid-19 Community Group Support Funding September 2020
CEF - Covid-19 Community Group Support Funding September 2020 Offering financial assistance to local volunteer groups and networks who are responding to and recovering from the Covid-19 crisis. Please note the funding should only be used for such associated costs. Aim of the funding The aim of The CEF Covid-19 community support funding is to provide a ‘community float’. This will enable local voluntary groups to access financial assistance to support their activities that help communities to connect again. How do community networks access the funding? The funding is held by the Community Support Organisations (CSO) to administer on behalf of Selby District Council. The CSO will be responsible for coordinating and supporting a network of local voluntary community groups. Under this agreement the CSO will agree a simple payment system with each group across the CSOs network can access the fund. Each group that receives funding will need to produce proof of expenditure (i.e. receipts/invoices.) What expenditure is eligible? The funding can be used flexibly to enable groups to operate smoothly without financial barriers. How the funding has been used will be reviewed / discussed at each reporting stage but it is reasonable to conclude that this is likely to be: . Volunteer expenses (might be mileage, refreshments) in relation to covid-19 response/recovery activity . PPE (gloves, sanitisers etc) . Cleaning/sanitary equipment/items . Administrative/marketing supplies (printing, stamps, stationary etc) . Equipment to enable delivery of services (i.e. phone credit, games, social distance signage) What is not eligible? . Personal payments or financial assistance of any kind. -
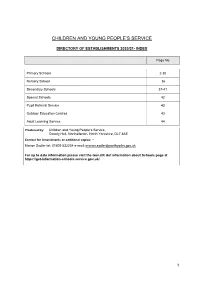
Directory of Establishments 2020/21- Index
CHILDREN AND YOUNG PEOPLE’S SERVICE DIRECTORY OF ESTABLISHMENTS 2020/21- INDEX Page No Primary Schools 2-35 Nursery School 36 Secondary Schools 37-41 Special Schools 42 Pupil Referral Service 43 Outdoor Education Centres 43 Adult Learning Service 44 Produced by: Children and Young People’s Service, County Hall, Northallerton, North Yorkshire, DL7 8AE Contact for Amendments or additional copies: – Marion Sadler tel: 01609 532234 e-mail: [email protected] For up to date information please visit the Gov.UK Get information about Schools page at https://get-information-schools.service.gov.uk/ 1 PRIMARY SCHOOLS Status Telephone County Council Ward School name and address Headteacher DfE No NC= nursery Email District Council area class Admiral Long Church of England Primary Mrs Elizabeth T: 01423 770185 3228 VC Lower Nidderdale & School, Burnt Yates, Harrogate, North Bedford E:admin@bishopthorntoncofe. Bishop Monkton Yorkshire, HG3 3EJ n-yorks.sch.uk Previously Bishop Thornton C of E Primary Harrogate Collaboration with Birstwith CE Primary School Ainderby Steeple Church of England Primary Mrs Fiona Sharp T: 01609 773519 3000 Academy Swale School, Station Lane, Morton On Swale, E: [email protected] Northallerton, North Yorkshire, Hambleton DL7 9QR Airy Hill Primary School, Waterstead Lane, Mrs Catherine T: 01947 602688 2190 Academy Whitby/Streonshalh Whitby, North Yorkshire, YO21 1PZ Mattewman E: [email protected] Scarborough NC Aiskew, Leeming Bar Church of England Mrs Bethany T: 01677 422403 3001 VC Swale Primary School, 2 Leeming Lane, Leeming Bar, Stanley E: admin@aiskewleemingbar. Northallerton, North Yorkshire, DL7 9AU n-yorks.sch.uk Hambleton Alanbrooke Community Primary School, Mrs Pippa Todd T: 01845 577474 2150 CS Sowerby Alanbrooke Barracks, Topcliffe, Thirsk, North E: admin@alanbrooke. -

Annual Review of Locality Budgets
North Yorkshire County Council Corporate and Partnerships Overview and Scrutiny Committee 7 June 2021 Annual review of County Councillor Locality Budgets 2020/21 1. Purpose of the report 1.1. To review the operation of County Councillor Locality Budgets in the financial year 2020/21. 2. Background 2.1. Locality Budgets enable County Councillors to respond to local needs by recommending the allocation of small amounts of funding to support projects or activities that directly promote the social, economic or environmental wellbeing of the communities they represent. 2.2. The Locality Budgets scheme seeks to ensure that County Councillors have a clear role in the allocation of funding in their Division within the criteria and process agreed by the Executive. This role includes actively promoting the scheme within their Division so that the money is allocated to meet appropriate local needs. 2.3. Because the allocation of funding is an executive decision, decisions are made by the Executive Member with responsibility for Stronger Communities, taking into account the information received and the recommendation of the relevant County Councillor. 2.4. Transparency is a significant feature of the scheme: County Councillors are required to declare all relevant interests when making a recommendation, including membership or other connections with organisations that will benefit from or manage a Locality Budget award Awards are published on the County Council website at https://www.northyorks.gov.uk/county-councillor-locality-budgets 1 OFFICIAL 3. Summary of approved recommendations in 2020/21 3.1. In 2020/21 each County Councillor had a Locality Budget allocation of £10,000. -

Appendix 2.4. Yorkshire Parliamentary Enclosure Awards
Changes in the status and distribution of mammals of the order Carnivora in Yorkshire from 1600. County history of the fox, badger, otter, pine marten, stoat, weasel, polecat, American mink, wildcat and domestic cat. Item Type Thesis Authors Howes, Colin Anthony Rights <a rel="license" href="http://creativecommons.org/licenses/ by-nc-nd/3.0/"><img alt="Creative Commons License" style="border-width:0" src="http://i.creativecommons.org/l/by- nc-nd/3.0/88x31.png" /></a><br />The University of Bradford theses are licenced under a <a rel="license" href="http:// creativecommons.org/licenses/by-nc-nd/3.0/">Creative Commons Licence</a>. Download date 25/09/2021 18:49:21 Link to Item http://hdl.handle.net/10454/4306 APPENDIX 2.4. YORKSHIRE PARLIAMENTARY ENCLOSURE AWARDS. APPENDIX 2.4. YORKSHIRE PARLIAMENTARY ENCLOSURE AWARDS (From English (1985) and Local Records Offices) (In alphabetical order of parish). Key: NR = North Riding, ER = East Riding, WR = West Riding, N = Nottinghamshire NY = North Yorkshire, SY = South Yorkshire, WY = West Yorkshire, H = Humberside, Cl = Cleveland, Cu = Cumbria , D = Durham, La = Lancashire, Li = Lincolnshire, Gm = Greater Manchester, Post Parish/Township Riding 1974 Act Award Acres County Abbotside, High NR NY 1814 1851 128 Abbotside, High & Low NR NY 1824 1837 938 Abbotside, High & Low NR NY 1880 1881 9701 Acklam ER NY 1769 1776 796 Acklam ER NY 1825 1854 310 Ackton WR WY 1812 1816 60 Ackworth WR WY 1772 1774 652 Acomb & Holgate WR NY 1774 1776 1581 Adingham WR WY 1865 1873 735 Adlingfleet WR H 1843 1847 1051 -

Superficial Deposits
Mineral Resource Information in Support of National, Regional and Local Planning: North Yorkshire (comprising North Yorkshire, Yorkshire Dales and North York Moors National Parks and City of York) Commissioned Report CR/04/228N BRITISH GEOLOGICAL SURVEY COMMISSIONED REPORT CR/04/228N Mineral Resource Information in Support of National, Regional and Local Planning: North Yorkshire (comprising North Yorkshire, Yorkshire Dales and North York Moors National Parks and City of York) D J Harrison, P J Henney, D Minchin, F M McEvoy, D G Cameron, S F Hobbs, D J Evans, G K Lott, E L Ball and D E Highley The National Grid and other Ordnance Survey data are used with the permission of the This report accompanies the two 1:100 000 scale maps: Controller of Her Majesty’s Stationery Office. North Yorkshire (comprising North Yorkshire, Yorkshire Ordnance Survey licence number GD 272191/2006 Dales and North York Moors National Parks and City of York). Key words North Yorkshire; mineral resources; mineral planning Front cover Coldstones Quarry, Carboniferous limestones, view to southwest. North Yorkshire Bibliographical reference Harrison, D J, Henney, P J, Minchin, D, McEvoy, F M, Cameron, D G, Hobbs, S F, Evans, D J, Lott, G K, Ball, E L, and Highley, D E. 2006. Mineral Resource Information in Support of National, Regional and Local Planning: North Yorkshire (comprising North Yorkshire, Yorkshire Dales and North York Moors National Parks and City of York). British Geological Survey Commissioned Report, CR/04/228N. 24pp © Crown Copyright 2006 Keyworth, Nottingham British Geological Survey 2006 BRITISH GEOLOGICAL SURVEY The full range of Survey publications is available Keyworth, Nottingham NG12 5GG from the BGS Sales Desks at Nottingham and Edinburgh; see contact details below or shop online 0115-936 3241 Fax 0115-936 at www.thebgs.co.uk 3488 e-mail: [email protected] The London Information Office maintains a www.bgs.ac.uk reference collection of BGS publications including Shop online at: www.thebgs.co.uk maps for consultation. -

Annual Report 2018/19
Annual Report 2018/19 Page 1 of 40 We are Citizens Advice and we are here for everyoneWe help people find a way forward We can all face problems that seem complicated or intimidating. At Citizens Advice Mid-North Yorkshire, we believe no one should have to face these problems without good quality, independent advice. That is why we are here: to give people the knowledge and the confidence they need to find their way forward - whoever they are, and whatever their problem. We give advice to thousands of people We are part of a network of independent charities that offers confidential advice online, over the phone, and in person, for free. Last year, Citizens Advice Mid-North Yorkshire helped 8,956 people with 36,923 issues, either face-to-face, telephone, web-chat or email. When we say we are for everyone, we mean it. People rely on us because we are independent and totally impartial. We are four services based at the very heart of our communities, in Hambleton, Richmondshire, Ryedale, and Selby & District. We speak up for our clients No one else sees so many people with so many different kinds of problems, and that gives us a unique insight into the challenges people are facing today. With the right evidence, we can show big organisations – from companies right up to the government – how they can make things better for people. We see how problems can be linked. By helping people with the underlying cause of their problems and making sure they don’t get worse; we save the government and public services hundreds of millions of pounds every year. -

W Om Er Sl Ey Parish M Agazine
YAGER COMPUTER SERVICES For help with all your IT problems W om er sl ey - Computer service & MOT - PC & Laptop repairs £30 per hour (excluding parts) - Upgrades Collection & delivery when possible. Par i sh M agazi n e - Home network installation For a friendly & reliable home service - Internet & WIFI help please contact - Virus & Spyware removal IAN YAGER - Home Computer security 07973 540946 - System setup and advise - Full assessment email:[email protected] - Clean-up & Optimisation web: www.yagerservices.co.uk STAPLETON DANCE & FITNESS STUDIO Bankwood Road, Stapleton, Pontefract, WF8 3DE Tel: 07900 856702 / 07789 084699 / 07729 191104 Email: [email protected] IDTA & UKA Registered ZUMBA FITNESS! MON 7.45pm ? 8.45pm / THURS 8.10pm ? 9.00pm --------------------------------------------------------- LEGS, BUMS & TUMS: MON 9:30am ? 10:30am / FRI 9:30am ? 10:30am DANCERCISE: MON 10:30am ? 11:30am "Be like a flower and turn your face to KEEP FIT: MON 12:15pm ? 1:15pm the sun." - Kahlil Gibran KETTLERCISE: MON 6:45pm ? 7:45pm BEGINNERS BALLROOM: TUES 7:30pm ? 8:30pm PILATES: TUES 7:30pm ? 8:30pm / WED 9:30am ? 10:30am PIYO: THURS 6:45pm ? 7:45pm SALSA CLASS: THURS 6:45pm ? 7:45pm M ay ADVANCED BALLROOM: FRI 7:30pm ? 9:00pm 20 17 KETTLEBELLS: SAT 9:30am ? 10:30am PRIVATE TUITION AVAILABLE 20 Ballroom , Lat in Am erican, Classical/ Modern Sequence, Salsa, Wedding Dances Ch u r ch of St M ar ti n Bu s an d Su p er m obi l e l i br ar y T i m etabl es Parish Priest Rev. Andrew Mart lew Arriva service No 409 - Doncast er t o Pont efract Services Monday t o Friday Family service every Sunday at 11.00am. -

Parish Registers Held in Local and Family History Complete List
Parish Registers held in Local and Family History Complete List PARISH REGISTERS IN LFH Page 2 of 49 Place Dates Format Where? Reel/Vol. Aberford 1540-1812 Printed L 906 THO (vol. 36) Alphabetical Aberford 1540-1885 Mic MIC 929.34274 Y82 1 Acklam 1716-1885 Mic MIC 929.34274 Y82 2 Ackworth 1558-1868 Mic MIC 929.34274 AC59 2 reels Ackworth 1813-1837 marriage index Printed FH ACK 929 Alphabetical Ackworth 1558-1600 Printed In Yorkshire Notes & Queries Ask staff (vol. 1) Y 052 Y82 Ackworth 1558-1957 Fiche Fiche Drawer Alphabetical Acomb 1634-1837 Printed Y 929.06 YOR 129 Acomb 1634-1885 Mic MIC 929.34274 Y82Y 3 Acomb 1725-1847 Mic MIC 929.34274 ACO Alphabetical Addingham 1612-1812 Printed Y 929.06 YOR 66 Addingham 1612-1959 Fiche Fiche Drawer Alphabetlcal Adel 1606-1812 Printed L 906 THO (vol. 5) Alphabetical Adel, St. John the Baptist 1606-2007 Fiche Fiche drawer Alphabetical Adwick-le-street 1547-1863 Mic MIC 929.34274 AD97 Alphabetical Airedale With Fryston 1905-1993 Fiche Fiche Drawer Alphabetical Aldborough 1538-1611 Printed Y 929.06 YOR 110 Aldborough 1538-1754 (name index only for Mic MIC 929.34274 Y82 9 1611-1629) Aldfield 1840 Mic Bishop’s Transcripts 116 Aldingham 1695-1756 Fiche MICF 929.06 LAN Misc. fiche Allerthorpe 1616-1885 Mic MIC 929.34274 Y82 3, 4 (seq. 1) Allerton, St Peter 1878-1990 Fiche Fiche Drawer Alphabetical Allerton Bywater, St. Mary the 1865-1980 Fiche Fiche drawer Alphabetical Less Allerton Mauleverer 1562-1966 Mic MIC 929.34274 AL54 Alphabetical Allerton Mauleverer 1674-1846 Mic Bishop’s Transcripts 116, 117 PARISH REGISTERS IN LFH Page 3 of 49 Place Dates Format Where? Reel/Vol. -

Inquisitions Post Mortem Relating to Yorkshire, of the Reigns of Henry IV
iiataljaU lEquttg Qlollcttton mn of IE. 3. MmaliM, ffi.ffi. 1. 1894 CORNELL UNIVERSITY LIBRARY 3 J924 084 250 624 u Cornell University Library The original of this book is in the Cornell University Library. There are no known copyright restrictions in the United States on the use of the text. http://www.archive.org/details/cu31924084250624 YORKSHIRE INQUISITIONS. VOL. V. THE YORKSHIRE ARCHAEOLOGICAL SOCIETY- Founded 1863. Incorporated 1893. RECORD SERIES, Vol. LIX. FOR THE YEAR 191 8. INQUISITIONS POST MORTEM RELATING TO YORKSHIRE, OF THE REIGNS OF HENRY IV AND HENRY V. KDITED BY W. PALEY BAILDON, F.S.A., AND J. W. CLAY, F.S.A. PRINTED FOR THE SOCIETY. 1918. PREFACE The present volume contains all the inquisitions post mortem, proofs of age and assignments of dower, relating to Yorkshire, for the reigns of Henry IV and Henry V, that are contained in the Chancery series. That series formerly included also the inquisitions ad quod damnum, which have now been made into a separate class, and are therefore not dealt with here. In view of the very full introduction to Vol. xii of the Record Series, it seems unnecessary to add to this volume any introduction on similar lines. The whole class of Chancery inquisitions post mortem is under arrangement; the documents are now arranged in files numbered from the beginning of each reign. The documents themselves, however, have not so far been renumbered, and still have the old system of numbering, beginning a new serial with each regnal year. It has therefore been thought better not to give the old serial number, in view of a probable renumbering at no distant date. -

Treasure Act Annual Report 2010 Presented to Parliament Pursuant to Section 12 of the Treasure Act 1996
Treasure Act Annual Report 2010 Presented to Parliament pursuant to Section 12 of the Treasure Act 1996 November 2012 Treasure Act Annual Report 2010 Presented to Parliament pursuant to Section 12 of the Treasure Act 1996 November 2012 i Minister’s foreword v Contents Introduction vii Treasure cases 2010 viii Tables of Treasure cases 2010 xiii Update on 2009 Treasure cases xlviii © Queen’s Printer and Controller of HMSO 2012 Edited by I Richardson, M Lewis, H Orange & C Lyons Published by the Department of Portable Antiquities and Treasure, British Museum ii iii Minister’s foreword It is my pleasure to introduce the Treasure Act Annual Report 2010. I would also like to acknowledge the valuable contribution of the Treasure Valuation Committee, The Treasure Act 1996 continues to be a great which provided valuation recommendations for success, ensuring that the most significant over 270 treasure cases from 2010. This summer archaeological discoveries are housed in public marks the first year of Professor Lord Renfrew of museums for the benefit of all. 2010 was a record Kaimsthorn’s Chairmanship of the Committee, and he year, with the number of potential Treasure finds has brought a wealth of archaeological knowledge reported increasing to 860 in England and Wales, and experience to the post. I am thankful for the a 10.5% increase since the previous year. Of these services of all of the Committee members, who items, 299 have been (or are to be) acquired by voluntarily provide their time and expertise. museums. It is especially satisfying that the number of interested parties waiving their right to a reward for Treasure finds remains high, with 86 individuals waiving their Ed Vaizey share in 70 cases.