Alan S. Weakley, Derick B. Poindexter, Hannah C. Medford
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Guide to the Flora of the Carolinas, Virginia, and Georgia, Working Draft of 17 March 2004 -- LILIACEAE
Guide to the Flora of the Carolinas, Virginia, and Georgia, Working Draft of 17 March 2004 -- LILIACEAE LILIACEAE de Jussieu 1789 (Lily Family) (also see AGAVACEAE, ALLIACEAE, ALSTROEMERIACEAE, AMARYLLIDACEAE, ASPARAGACEAE, COLCHICACEAE, HEMEROCALLIDACEAE, HOSTACEAE, HYACINTHACEAE, HYPOXIDACEAE, MELANTHIACEAE, NARTHECIACEAE, RUSCACEAE, SMILACACEAE, THEMIDACEAE, TOFIELDIACEAE) As here interpreted narrowly, the Liliaceae constitutes about 11 genera and 550 species, of the Northern Hemisphere. There has been much recent investigation and re-interpretation of evidence regarding the upper-level taxonomy of the Liliales, with strong suggestions that the broad Liliaceae recognized by Cronquist (1981) is artificial and polyphyletic. Cronquist (1993) himself concurs, at least to a degree: "we still await a comprehensive reorganization of the lilies into several families more comparable to other recognized families of angiosperms." Dahlgren & Clifford (1982) and Dahlgren, Clifford, & Yeo (1985) synthesized an early phase in the modern revolution of monocot taxonomy. Since then, additional research, especially molecular (Duvall et al. 1993, Chase et al. 1993, Bogler & Simpson 1995, and many others), has strongly validated the general lines (and many details) of Dahlgren's arrangement. The most recent synthesis (Kubitzki 1998a) is followed as the basis for familial and generic taxonomy of the lilies and their relatives (see summary below). References: Angiosperm Phylogeny Group (1998, 2003); Tamura in Kubitzki (1998a). Our “liliaceous” genera (members of orders placed in the Lilianae) are therefore divided as shown below, largely following Kubitzki (1998a) and some more recent molecular analyses. ALISMATALES TOFIELDIACEAE: Pleea, Tofieldia. LILIALES ALSTROEMERIACEAE: Alstroemeria COLCHICACEAE: Colchicum, Uvularia. LILIACEAE: Clintonia, Erythronium, Lilium, Medeola, Prosartes, Streptopus, Tricyrtis, Tulipa. MELANTHIACEAE: Amianthium, Anticlea, Chamaelirium, Helonias, Melanthium, Schoenocaulon, Stenanthium, Veratrum, Toxicoscordion, Trillium, Xerophyllum, Zigadenus. -
Species List (PDF)
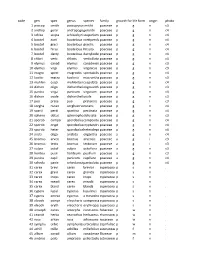
code gen spec genus species family growth formlife form origin photo 1 pascop smith pascopyrumsmithii poaceae p g n c3 2 androp gerar andropogongerardii poaceae p g n c4 3 schiza scopa schizachyriumscoparium poaceae p g n c4 4 boutel curti bouteloua curtipendulapoaceae p g n c4 5 boutel graci bouteloua gracilis poaceae p g n c4 6 boutel hirsu bouteloua hirsuta poaceae p g n c4 7 boutel dacty bouteloua dactyloidespoaceae p g n c4 8 chlori verti chloris verticillata poaceae p g n c4 9 elymus canad elymus canadensispoaceae p g n c3 10 elymus virgi elymus virginicus poaceae p g n c3 11 eragro spect eragrostis spectabilis poaceae p g n c4 12 koeler macra koeleria macrantha poaceae p g n c3 13 muhlen cuspi muhlenbergiacuspidata poaceae p g n c4 14 dichan oligo dichantheliumoligosanthespoaceae p g n c3 15 panicu virga panicum virgatum poaceae p g n c4 16 dichan ovale dichantheliumovale poaceae p g n c3 17 poa prate poa pratensis poaceae p g i c3 18 sorgha nutan sorghastrumnutans poaceae p g n c4 19 sparti pecti spartina pectinata poaceae p g n c4 20 spheno obtus sphenopholisobtusata poaceae p g n c3 21 sporob compo sporoboluscomposituspoaceae p g n c4 22 sporob crypt sporoboluscryptandruspoaceae p g n c4 23 sporob heter sporobolusheterolepispoaceae p g n c4 24 aristi oliga aristida oligantha poaceae a g n c4 25 bromus arven bromus arvensis poaceae a g i c3 26 bromus tecto bromus tectorum poaceae a g i c3 27 vulpia octof vulpia octoflora poaceae a g n c3 28 hordeu pusil hordeum pusillum poaceae a g n c3 29 panicu capil panicum capillare poaceae a g n c4 30 schedo panic schedonnarduspaniculatuspoaceae p g n c4 31 carex brevi carex brevior cyperaceaep s n . -

Pima County Plant List (2020) Common Name Exotic? Source
Pima County Plant List (2020) Common Name Exotic? Source McLaughlin, S. (1992); Van Abies concolor var. concolor White fir Devender, T. R. (2005) McLaughlin, S. (1992); Van Abies lasiocarpa var. arizonica Corkbark fir Devender, T. R. (2005) Abronia villosa Hariy sand verbena McLaughlin, S. (1992) McLaughlin, S. (1992); Van Abutilon abutiloides Shrubby Indian mallow Devender, T. R. (2005) Abutilon berlandieri Berlandier Indian mallow McLaughlin, S. (1992) Abutilon incanum Indian mallow McLaughlin, S. (1992) McLaughlin, S. (1992); Van Abutilon malacum Yellow Indian mallow Devender, T. R. (2005) Abutilon mollicomum Sonoran Indian mallow McLaughlin, S. (1992) Abutilon palmeri Palmer Indian mallow McLaughlin, S. (1992) Abutilon parishii Pima Indian mallow McLaughlin, S. (1992) McLaughlin, S. (1992); UA Abutilon parvulum Dwarf Indian mallow Herbarium; ASU Vascular Plant Herbarium Abutilon pringlei McLaughlin, S. (1992) McLaughlin, S. (1992); UA Abutilon reventum Yellow flower Indian mallow Herbarium; ASU Vascular Plant Herbarium McLaughlin, S. (1992); Van Acacia angustissima Whiteball acacia Devender, T. R. (2005); DBGH McLaughlin, S. (1992); Van Acacia constricta Whitethorn acacia Devender, T. R. (2005) McLaughlin, S. (1992); Van Acacia greggii Catclaw acacia Devender, T. R. (2005) Acacia millefolia Santa Rita acacia McLaughlin, S. (1992) McLaughlin, S. (1992); Van Acacia neovernicosa Chihuahuan whitethorn acacia Devender, T. R. (2005) McLaughlin, S. (1992); UA Acalypha lindheimeri Shrubby copperleaf Herbarium Acalypha neomexicana New Mexico copperleaf McLaughlin, S. (1992); DBGH Acalypha ostryaefolia McLaughlin, S. (1992) Acalypha pringlei McLaughlin, S. (1992) Acamptopappus McLaughlin, S. (1992); UA Rayless goldenhead sphaerocephalus Herbarium Acer glabrum Douglas maple McLaughlin, S. (1992); DBGH Acer grandidentatum Sugar maple McLaughlin, S. (1992); DBGH Acer negundo Ashleaf maple McLaughlin, S. -

SFRC T-593 Phenology of Flowering and Fruiting
Report T-593 Phenology of Flowering an Fruiting In Pia t Com unities of Everglades NP and Biscayne N , orida RESOURCE MANAGEMENT EVERGLi\DES NATIONAL PARK BOX 279 NOMESTEAD, FLORIDA 33030 Everglades National Park, South Florida Research Center, P.O. Box 279, Homestead, Florida 33030 PHENOLOGY OF FLOWERING AND FRUITING IN PLANT COMMUNITIES OF EVERGLADES NATIONAL PARK AND BISCAYNE NATIONAL MONUMENT, FLORIDA Report T - 593 Lloyd L. Loope U.S. National P ark Service South Florida Research Center Everglades National Park Homestead, Florida 33030 June 1980 Loope, Lloyd L. 1980. Phenology of Flowering and Fruiting in Plant Communities of Everglades National Park and Biscayne National Monument, Florida. South Florida Research Center Report T - 593. 50 pp. TABLE OF CONTENTS LIST OF TABLES • ii LIST OF FIGU RES iv INTRODUCTION • 1 ACKNOWLEDGEMENTS. • 1 METHODS. • • • • • • • 1 CLIMATE AND WATER LEVELS FOR 1978 •• . 3 RESULTS ••• 3 DISCUSSION. 3 The need and mechanisms for synchronization of reproductive activity . 3 Tropical hardwood forest. • • 5 Freshwater wetlands 5 Mangrove vegetation 6 Successional vegetation on abandoned farmland. • 6 Miami Rock Ridge pineland. 7 SUMMARY ••••• 7 LITERATURE CITED 8 i LIST OF TABLES Table 1. Climatic data for Homestead Experiment Station, 1978 . • . • . • . • . • . • . 10 Table 2. Climatic data for Tamiami Trail at 40-Mile Bend, 1978 11 Table 3. Climatic data for Flamingo, 1978. • • • • • • • • • 12 Table 4. Flowering and fruiting phenology, tropical hardwood hammock, area of Elliott Key Marina, Biscayne National Monument, 1978 • • • • • • • • • • • • • • • • • • 14 Table 5. Flowering and fruiting phenology, tropical hardwood hammock, Bear Lake Trail, Everglades National Park (ENP), 1978 • . • . • . 17 Table 6. Flowering and fruiting phenology, tropical hardwood hammock, Mahogany Hammock, ENP, 1978. -

Outline of Angiosperm Phylogeny
Outline of angiosperm phylogeny: orders, families, and representative genera with emphasis on Oregon native plants Priscilla Spears December 2013 The following listing gives an introduction to the phylogenetic classification of the flowering plants that has emerged in recent decades, and which is based on nucleic acid sequences as well as morphological and developmental data. This listing emphasizes temperate families of the Northern Hemisphere and is meant as an overview with examples of Oregon native plants. It includes many exotic genera that are grown in Oregon as ornamentals plus other plants of interest worldwide. The genera that are Oregon natives are printed in a blue font. Genera that are exotics are shown in black, however genera in blue may also contain non-native species. Names separated by a slash are alternatives or else the nomenclature is in flux. When several genera have the same common name, the names are separated by commas. The order of the family names is from the linear listing of families in the APG III report. For further information, see the references on the last page. Basal Angiosperms (ANITA grade) Amborellales Amborellaceae, sole family, the earliest branch of flowering plants, a shrub native to New Caledonia – Amborella Nymphaeales Hydatellaceae – aquatics from Australasia, previously classified as a grass Cabombaceae (water shield – Brasenia, fanwort – Cabomba) Nymphaeaceae (water lilies – Nymphaea; pond lilies – Nuphar) Austrobaileyales Schisandraceae (wild sarsaparilla, star vine – Schisandra; Japanese -

Chamaecrista Fasciculata in Tallgrass and Sand Prairies: the Potential for Differential Responses
Illinois State University ISU ReD: Research and eData Theses and Dissertations 6-27-2018 Chamaecrista Fasciculata In Tallgrass And Sand Prairies: The Potential For Differential Responses Robert W. Philips Illinois State University, [email protected] Follow this and additional works at: https://ir.library.illinoisstate.edu/etd Part of the Botany Commons, and the Evolution Commons Recommended Citation Philips, Robert W., "Chamaecrista Fasciculata In Tallgrass And Sand Prairies: The Potential For Differential Responses" (2018). Theses and Dissertations. 918. https://ir.library.illinoisstate.edu/etd/918 This Thesis is brought to you for free and open access by ISU ReD: Research and eData. It has been accepted for inclusion in Theses and Dissertations by an authorized administrator of ISU ReD: Research and eData. For more information, please contact [email protected]. CHAMAECRISTA FASCICULATA IN TALLGRASS AND SAND PRAIRIES: THE POTENTIAL FOR DIFFERENTIAL RESPONSES ROBERT W. PHILIPS 90 Pages Successful establishment of a diversity of native species has become an important goal for restoration site managers to achieve, however as seed sources for a species may occur in habitats with different abiotic and biotic characteristics. Consequently, seeds from different sources may vary in their success in a restoration. Chamaecrista fasciculata, a native prairie species, occurs in two divergent prairie types - tallgrass and sand prairies. Tallgrass prairies have a moist soil with dense vegetation; in contrast, sand prairies have a well-drained sandy soil with sparse vegetation. I propose differential selection acting on populations in these prairie types would affect their seeds success in restorations. Given the denser vegetation of the tallgrass prairies, plants must be capable of competing for light resources, thus I predict the plants from tallgrass seed sources have a better competitor tolerance and would be more successful in a reconstructed tallgrass prairie. -

State of New York City's Plants 2018
STATE OF NEW YORK CITY’S PLANTS 2018 Daniel Atha & Brian Boom © 2018 The New York Botanical Garden All rights reserved ISBN 978-0-89327-955-4 Center for Conservation Strategy The New York Botanical Garden 2900 Southern Boulevard Bronx, NY 10458 All photos NYBG staff Citation: Atha, D. and B. Boom. 2018. State of New York City’s Plants 2018. Center for Conservation Strategy. The New York Botanical Garden, Bronx, NY. 132 pp. STATE OF NEW YORK CITY’S PLANTS 2018 4 EXECUTIVE SUMMARY 6 INTRODUCTION 10 DOCUMENTING THE CITY’S PLANTS 10 The Flora of New York City 11 Rare Species 14 Focus on Specific Area 16 Botanical Spectacle: Summer Snow 18 CITIZEN SCIENCE 20 THREATS TO THE CITY’S PLANTS 24 NEW YORK STATE PROHIBITED AND REGULATED INVASIVE SPECIES FOUND IN NEW YORK CITY 26 LOOKING AHEAD 27 CONTRIBUTORS AND ACKNOWLEGMENTS 30 LITERATURE CITED 31 APPENDIX Checklist of the Spontaneous Vascular Plants of New York City 32 Ferns and Fern Allies 35 Gymnosperms 36 Nymphaeales and Magnoliids 37 Monocots 67 Dicots 3 EXECUTIVE SUMMARY This report, State of New York City’s Plants 2018, is the first rankings of rare, threatened, endangered, and extinct species of what is envisioned by the Center for Conservation Strategy known from New York City, and based on this compilation of The New York Botanical Garden as annual updates thirteen percent of the City’s flora is imperiled or extinct in New summarizing the status of the spontaneous plant species of the York City. five boroughs of New York City. This year’s report deals with the City’s vascular plants (ferns and fern allies, gymnosperms, We have begun the process of assessing conservation status and flowering plants), but in the future it is planned to phase in at the local level for all species. -

Hazardous Fuels Management in Subtropical Pine Flatwoods and Tropical Pine Rocklands Joseph J
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln JFSP Research Project Reports U.S. Joint Fire Science Program 2007 Fire Managers Field Guide: Hazardous Fuels Management in Subtropical Pine Flatwoods and Tropical Pine Rocklands Joseph J. O'Brien USDA Forest Service Kathyryn A. Mordecai USDA Forest Service Leslie Wolcott USDA Forest Service James Snyder USGS Kenneth Outcalt USDA Forest Service Follow this and additional works at: http://digitalcommons.unl.edu/jfspresearch Part of the Forest Biology Commons, Forest Management Commons, Natural Resources and Conservation Commons, Natural Resources Management and Policy Commons, Other Environmental Sciences Commons, Other Forestry and Forest Sciences Commons, Sustainability Commons, and the Wood Science and Pulp, Paper Technology Commons O'Brien, Joseph J.; Mordecai, Kathyryn A.; Wolcott, Leslie; Snyder, James; and Outcalt, Kenneth, "Fire Managers Field Guide: Hazardous Fuels Management in Subtropical Pine Flatwoods and Tropical Pine Rocklands" (2007). JFSP Research Project Reports. 101. http://digitalcommons.unl.edu/jfspresearch/101 This Article is brought to you for free and open access by the U.S. Joint Fire Science Program at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in JFSP Research Project Reports by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. Fire Managers Field Guide: Hazardous Fuels Management in Subtropical Pine Flatwoods and Tropical Pine Rocklands FFFiiinnnaaalll RRReeepppooorrrttt -

Carex Gigantea Rudge in Florida
ILLINOI S UNIVERSITY OF ILLINOIS AT URBANA-CHAMPAIGN PRODUCTION NOTE University of Illinois at Urbana-Champaign Library Large-scale Digitization Project, 2007. c f ConservationAssessment for the Giant Sedge (Carex giganteaRudge) 31 July 2006 Steven R. Hill, Ph.D. Illinois Natural History Survey Center for Wildlife and Plant Ecology 1816 South Oak Street Champaign, Illinois 61820 ILLINOIS NATURAL HISTORY SURVEY Illinois Natural History Survey Center for Wildlife and Plant Ecology Technical Report 2006 (10) Cover photo: Carex gigantea Rudge in Florida. Photo by Shirley Denton. http://www.simple-grandeur.com/plants/native.php This Conservation Assessment was prepared to compile the published and unpublished information on the subject taxon or community; or this document was prepared by another organization and provides information to serve as a Conservation Assessment for the Eastern Region of the Forest Service. It does not represent a management decision by the U.S. Forest Service. Though the best scientific information available was used and subject experts were consulted in preparation of this document, it is expected that new information will arise. In the spirit of continuous learning and adaptive management, if you have information that will assist in conserving the subject taxon, please contact the Eastern Region of the Forest Service - Threatened and Endangered Species Program at 310 Wisconsin Avenue, Suite 580 Milwaukee, Wisconsin 53203. 2 Conservation Assessment for the Giant Sedge (Carex gigantea Rudge) Table of Contents Acknowledgm -
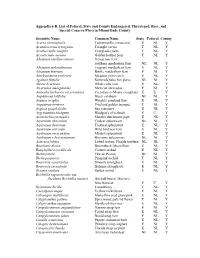
Conservation Appendix 6-B Listed Flora
Appendix 6-B. List of Federal, State and County Endangered, Threatened, Rare, and Special Concern Flora in Miami-Dade County Scientific Name Common Name State Federal County Acacia choriophylla Tamarindillo; cinnecord E NL Y Acanthocereus tetragenus Triangle cactus T NL Y Acoelorraphe wrightii Everglades palm T NL Y Acrostichum aureum Golden leather fern T NL Y Adiantum capillus-veneris Venus hair fern; southern maidenhair fern NL NL Y Adiantum melanoleucum Fragrant maidenhair fern E NL Y Adiantum tenerum Brittle maidenhair fern E NL Y Aeschynomene pratensis Meadow joint-vetch E NL Y Agalinis filifolia Seminole false fox glove NL NL Y Aletris bracteata White colic root E NL Y Alvaradoa amorphoides Mexican alvaradoa E NL Y Amorpha herbacea var.crenulata Crenulate (=Miami) leadplant E E Y Amphitecna latifolia Black calabash NL NL Y Anemia wrightii Wright's pineland fern E NL Y Angadenia berteroi Pineland golden trumpet T NL Y Argusia gnaphalodes Sea rosemary E NL Y Argythamnia blodgettii Blodgett's silverbush E C Y Aristolochia pentandra Marsh's dutchmans pipe E NL Y Asplenium abscissum Cutleaf spleenwort NL NL Y Asplenium dentatum Toothed spleenwort E NL Y Asplenium serratum Wild bird nest fern E NL Y Asplenium verecundum Modest spleenwort E NL Y Asplenium x biscaynianum Biscayne spleenwort NL NL Y Asteraea lobata Lobed croton; Florida treefern NL NL Y Baccharis dioica Broombush falsewillow E NL Y Basiphyllaea corallicola Carter's orchid E NL Y Bletia patula Flor de Pesmo NL NL Y Bletia purpurea Pinepink orchid T NL Y Bourreria cassinifolia Smooth strongback E NL Y Bourreria succulenta Bahama strongback E NL Y Brassia caudata Spider orchid E NL Y Brickellia eupatorioides var. -

Genetic Diversity of Chamaecrista Fasciculata (Fabaceae) from the USDA Germplasm Collection
University of Vermont ScholarWorks @ UVM College of Agriculture and Life Sciences Faculty Publications College of Agriculture and Life Sciences 3-4-2019 Genetic diversity of Chamaecrista fasciculata (Fabaceae) from the USDA germplasm collection Erika Bueno University of Vermont Ted Kisha USDA Agricultural Research Service, Washington DC Sonja L. Maki Carleton College, USA Eric J.B. Von Wettberg University of Vermont Susan Singer Carleton College, USA Follow this and additional works at: https://scholarworks.uvm.edu/calsfac Part of the Community Health Commons, Human Ecology Commons, Nature and Society Relations Commons, Place and Environment Commons, and the Sustainability Commons Recommended Citation Bueno E, Kisha T, Maki SL, von Wettberg EJ, Singer S. Genetic diversity of Chamaecrista fasciculata (Fabaceae) from the USDA germplasm collection. BMC research notes. 2019 Dec 1;12(1):117. This Article is brought to you for free and open access by the College of Agriculture and Life Sciences at ScholarWorks @ UVM. It has been accepted for inclusion in College of Agriculture and Life Sciences Faculty Publications by an authorized administrator of ScholarWorks @ UVM. For more information, please contact [email protected]. Bueno et al. BMC Res Notes (2019) 12:117 https://doi.org/10.1186/s13104-019-4152-0 BMC Research Notes RESEARCH NOTE Open Access Genetic diversity of Chamaecrista fasciculata (Fabaceae) from the USDA germplasm collection Erika Bueno1, Ted Kisha2, Sonja L. Maki3,4, Eric J. B. von Wettberg1* and Susan Singer3,5 Abstract Objective: Chamaecrista fasciculata is a widespread annual legume across Eastern North America, with potential as a restoration planting, biofuel crop, and genetic model for non-papillinoid legumes. -

BC-TES-5759.Pdf (1.465Mb)
UNIVERSIDAD NACIONAL “PEDRO RUIZ GALLO” FACULTAD DE CIENCIAS BIOLÓGICAS DEPARTAMENTO ACADEMICO DE MICROBIOLOGIA Y PARASITOLOGÍA ACTIVIDAD ANTIBACTERIANA, In Vitro DEL EXTRACTO ETANOLICO DE Morinda citrifolia L. “Noni” FRENTE A CEPAS DE Pseudomonas aeruginosa y Staphylococcus aureus. TESIS: PARA OPTAR EL TITULO PROFESIONAL DE LICENCIADO EN BIOLOGIA – MICROBIOLOGIA Y PARASITOLOGIA PRESENTADO POR: Br. Luis Ángel Altamirano Fernández. Br. Emeli Maday Castro Bruno. LAMBAYEQUE – PERU 2017 UNIVERSIDAD NACIONAL “PEDRO RUIZ GALLO” FACULTAD DE CIENCIAS BIOLÓGICAS DEPARTAMENTO ACADEMICO DE MICROBIOLOGIA Y PARASITOLOGÍA ACTIVIDAD ANTIBACTERIANA, In Vitro DEL EXTRACTO ETANOLICO DE Morinda citrifolia L. “Noni” FRENTE A CEPAS DE Pseudomonas aeruginosa y Staphylococcus aureus. TESIS PARA OPTAR EL TITULO PROFESIONAL DE LICENCIADO EN: BIOLOGIA – MICROBIOLOGIA Y PARASITOLOGIA AUTORES: Br. Luis Ángel Altamirano Fernández. Br. Emeli Maday Castro Bruno. LAMBAYEQUE - PERU 2017 UNIVERSIDAD NACIONAL “PEDRO RUIZ GALLO” FACULTAD DE CIENCIAS BIOLÓGICAS DEPARTAMENTO ACADEMICO DE MICROBIOLOGIA Y PARASITOLOGÍA ACTIVIDAD ANTIBACTERIANA, In Vitro DEL EXTRACTO ETANOLICO DE Morinda citrifolia L. “Noni” FRENTE A CEPAS DE Pseudomonas aeruginosa y Staphylococcus aureus. Luis Ángel Altamirano Fernández. Emeli Maday Castro Bruno. TESIS PRESENTADA PARA OPTAR EL TITULO DE LICENCIADO EN: BIOLOGIA – MICROBIOLOGIA Y PARASITOLOGIA APROBADA POR: Mblga. María Teresa Silva García PRESIDENTE Dra. Gianina Llontop Barandiaran SECRETARIA Lic. Julio Cesar Silva Estela VOCAL Lic. Mario Cecilio Moreno Mantilla ASESOR Lic. Fransk Amarildo Carrasco Solano CO-ASESOR Aquel que obtiene una victoria sobre otro hombre es fuerte, pero quien obtiene una victoria sobre sí mismo es poderoso. LAO TZE DEDICATORIAS Dedico mi tesis con todo mi corazón y cariño a mi padre Félix Altamirano Campos, que trabajó arduamente para poder darme una carrera universitaria, por su apoyo incondicional y su cariño, sé que desde el cielo está orgulloso de mí y me da fuerzas para seguir superándome.