Chamaecyparis Montane Cloud Forest in Taiwan: Ecology and Vegetation Classification
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Department of Planning and Zoning
Department of Planning and Zoning Subject: Howard County Landscape Manual Updates: Recommended Street Tree List (Appendix B) and Recommended Plant List (Appendix C) - Effective July 1, 2010 To: DLD Review Staff Homebuilders Committee From: Kent Sheubrooks, Acting Chief Division of Land Development Date: July 1, 2010 Purpose: The purpose of this policy memorandum is to update the Recommended Plant Lists presently contained in the Landscape Manual. The plant lists were created for the first edition of the Manual in 1993 before information was available about invasive qualities of certain recommended plants contained in those lists (Norway Maple, Bradford Pear, etc.). Additionally, diseases and pests have made some other plants undesirable (Ash, Austrian Pine, etc.). The Howard County General Plan 2000 and subsequent environmental and community planning publications such as the Route 1 and Route 40 Manuals and the Green Neighborhood Design Guidelines have promoted the desirability of using native plants in landscape plantings. Therefore, this policy seeks to update the Recommended Plant Lists by identifying invasive plant species and disease or pest ridden plants for their removal and prohibition from further planting in Howard County and to add other available native plants which have desirable characteristics for street tree or general landscape use for inclusion on the Recommended Plant Lists. Please note that a comprehensive review of the street tree and landscape tree lists were conducted for the purpose of this update, however, only -

Morphology and Morphogenesis of the Seed Cones of the Cupressaceae - Part II Cupressoideae
1 2 Bull. CCP 4 (2): 51-78. (10.2015) A. Jagel & V.M. Dörken Morphology and morphogenesis of the seed cones of the Cupressaceae - part II Cupressoideae Summary The cone morphology of the Cupressoideae genera Calocedrus, Thuja, Thujopsis, Chamaecyparis, Fokienia, Platycladus, Microbiota, Tetraclinis, Cupressus and Juniperus are presented in young stages, at pollination time as well as at maturity. Typical cone diagrams were drawn for each genus. In contrast to the taxodiaceous Cupressaceae, in Cupressoideae outgrowths of the seed-scale do not exist; the seed scale is completely reduced to the ovules, inserted in the axil of the cone scale. The cone scale represents the bract scale and is not a bract- /seed scale complex as is often postulated. Especially within the strongly derived groups of the Cupressoideae an increased number of ovules and the appearance of more than one row of ovules occurs. The ovules in a row develop centripetally. Each row represents one of ascending accessory shoots. Within a cone the ovules develop from proximal to distal. Within the Cupressoideae a distinct tendency can be observed shifting the fertile zone in distal parts of the cone by reducing sterile elements. In some of the most derived taxa the ovules are no longer (only) inserted axillary, but (additionally) terminal at the end of the cone axis or they alternate to the terminal cone scales (Microbiota, Tetraclinis, Juniperus). Such non-axillary ovules could be regarded as derived from axillary ones (Microbiota) or they develop directly from the apical meristem and represent elements of a terminal short-shoot (Tetraclinis, Juniperus). -

Phylogeny and Biogeography of Tsuga (Pinaceae)
Systematic Botany (2008), 33(3): pp. 478–489 © Copyright 2008 by the American Society of Plant Taxonomists Phylogeny and Biogeography of Tsuga (Pinaceae) Inferred from Nuclear Ribosomal ITS and Chloroplast DNA Sequence Data Nathan P. Havill1,6, Christopher S. Campbell2, Thomas F. Vining2,5, Ben LePage3, Randall J. Bayer4, and Michael J. Donoghue1 1Department of Ecology and Evolutionary Biology, Yale University, New Haven, Connecticut 06520-8106 U.S.A 2School of Biology and Ecology, University of Maine, Orono, Maine 04469-5735 U.S.A. 3The Academy of Natural Sciences, 1900 Benjamin Franklin Parkway, Philadelphia, Pennsylvania 19103 U.S.A. 4CSIRO – Division of Plant Industry, Center for Plant Biodiversity Research, GPO 1600, Canberra, ACT 2601 Australia; present address: Department of Biology, University of Memphis, Memphis, Tennesee 38152 U.S.A. 5Present address: Delta Institute of Natural History, 219 Dead River Road, Bowdoin, Maine 04287 U.S.A. 6Author for correspondence ([email protected]) Communicating Editor: Matt Lavin Abstract—Hemlock, Tsuga (Pinaceae), has a disjunct distribution in North America and Asia. To examine the biogeographic history of Tsuga, phylogenetic relationships among multiple accessions of all nine species were inferred using chloroplast DNA sequences and multiple cloned sequences of the nuclear ribosomal ITS region. Analysis of chloroplast and ITS sequences resolve a clade that includes the two western North American species, T. heterophylla and T. mertensiana, and a clade of Asian species within which one of the eastern North American species, T. caroliniana, is nested. The other eastern North American species, T. canadensis, is sister to the Asian clade. Tsuga chinensis from Taiwan did not group with T. -

Nursery Catalog
Tel: 503.628.8685 Fax: 503.628.1426 www.eshraghinursery.com 1 Eshraghi’s TOP 10 picks Our locations 1 Main Office, Shipping & Growing 2 Retail Store & Growing 26985 SW Farmington Road Farmington Gardens Hillsboro, OR 97123 21815 SW Farmington Road Beaverton, OR 97007 1 2 3 7 6 3 River Ranch Facility 4 Liberty Farm 4 5 10 N SUNSET HWY TO PORTLAND 8 9 TU HILLSBORO ALA TIN 26 VALL SW 185TH AVE. EY HWY. #4 8 BEAVERTON TONGUE LN. GRABEL RD . D R . E D G R ID E ALOHA R G B D I R R #3 SW 209TH E B T D FARMINGTON ROAD D N A I SIMPSON O O M O R R 10 217 ROSEDALE W R E S W V S I R N W O 219 T K C A J #2 #1 SW UNGER RD. SW 185TH AVE. 1 Acer circinatum ‘Pacific Fire’ (Vine Maple), page 6 D A SW MURRAY BLVD. N RO 2 palmatum (Japanese Maple), NGTO Acer 'Geisha Gone Wild' page 8 FARMI 3 Acer palmatum 'Mikawa yatsubusa' (Japanese Maple), page 10 #1 4 Acer palmatum dissectum 'Orangeola' (Japanese Maple), page 14 5 Hydrangea macrophylla 'McKay', Cherry Explosion PP28757 (Hydrangea), page 32 6 Picea glauca 'Eshraghi1', Poco Verde (White Spruce), page 61 ROAD HILL CLARK 7 Picea pungens 'Hockersmith', Linda (Colorado Spruce), page 64 RY ROAD 8 Pinus nigra 'Green Tower' (Austrian Pine), page 65 SCHOLLS FER 9 Thuja occidentalis 'Janed Gold', Highlights™ PP21967 (Arborvitae), page 70 10 Thuja occidentalis 'Anniek', Sienna Sunset™ (Arborvitae), page 69 Table of contents Tags Make a Difference . -

The Baker's Cypress
AMERICAN CONIFER SOCIETY coniferVOLUME 33, NUMBER 2 | SPRING 2016 QUARTERLY ENCOUNTERS WITH The Baker’s Cypress PAGE 18 SAVE THE DATE • 2016 SOUTHEAST REGION MEETING • AUGUST 26–28 • WAYNESBORO, VA TABLE O F CONTENTS 16 05 18 12 Welcome to the new ConiferQuarterly ACS Seed Exchange and How I Became By Ron Elardo 04 16 a Coniferite By Jim Brackman What Do Conifer Enthusiasts Need to Encounters with The Baker’s Cypress Know About Mycorrhizae? 05 18 By David Pilz By Bert Cregg, Ph.D. Comments on Conifers for Open Forum: Southeast Region ACS Part 1 09 22 Reference Gardens By Bob Fincham 2016 Southeast Region Meeting ACS Directorate By Jeff Harvey 12 23 Shady Characters: Conifers and Plants Made For Shade 14 By Rich and Susan Eyre Spring 2016 Volume 33, Number 2 ConiferQuarterly (ISSN 8755-0490) is published quarterly by the American Conifer Society. The Society is a non- Conifer profit organization incorporated under the laws of the Commonwealth of Pennsylvania and is tax exempt under Quarterly section 501(c)3 of the Internal Revenue Service Code. You are invited to join our Society. Please address Editor membership and other inquiries to the American Conifer Ronald J. Elardo Society National Office, PO Box 1583, Minneapolis, MN 55311, [email protected]. Membership: US & Canada $38, International $58 (indiv.), $30 (institutional), $50 Technical Editors (sustaining), $100 (corporate business) and $130 (patron). Steven Courtney If you are moving, please notify the National Office 4 weeks Robert Fincham in advance. Ethan Johnson David Olszyk All editorial and advertising matters should be sent to: Ron Elardo, 5749 Hunter Ct., Adrian, MI 49221-2471, (517) 902-7230 or email [email protected] Advisory Committee Tom Neff, Committee Chair Copyright © 2016, American Conifer Society. -
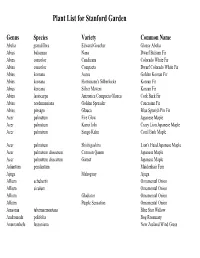
Plant List for Stanford Garden
Plant List for Stanford Garden Genus Species Variety Common Name Abelia grandiflora Edward Goucher Glossy Abelia Abies balsamea Nana Dwarf Balsam Fir Abies concolor Candicans Colorado White Fir Abies concolor Compacta Dwarf Colorado White Fir Abies koreana Aurea Golden Korean Fir Abies koreana Hortsmann's Silberlocke Korean Fir Abies koreana Silber Mavers Korean Fir Abies lasiocarpa Arizonica Compacta Glauca Cork Bark Fir Abies nordmanniana Golden Spreader Caucasian Fir Abies pinsapo Glauca Blue Spanish Pin Fir Acer palmatum Fire Glow Japanese Maple Acer palmatum Kurui Jishi Crazy Lion Japanese Maple Acer palmatum Sango Kaku Coral Bark Maple Acer palmatum Shishigashira Lion's Head Japanese Maple Acer palmatum dissectum Crimson Queen Japanese Maple Acer palmatum dissectum Garnet Japanese Maple Adiantum pendantum Maidenhair Fern Ajuga Mahogany Ajuga Allium schubertii Ornamental Onion Allium siculum Ornamental Onion Allium Gladiator Ornamental Onion Allium Purple Sensation Ornamental Onion Amsonia tabernaemontana Blue Star Willow Andromeda polifolia Bog Rosemary Anemanthele lessoniana New Zealand Wind Grass Genus Species Variety Common Name Anemone coronaria de Caen Wind Flower Anemone hupehensis japonica September Charm Japanese Anemone Anemone huphensis japonica Pamina Japanese Anemone Anemone x hybrida Honorine Jobert Japanese Anemone Angelica Aquilegia vulgaris Lime Frost Columbine Aquilegia Woodside Golden Columbine Aquillega Yellow Queen Yellow Columbine Arabis caucasica Variegata Variegated Rock Cress Arisaema formosanum DJHT 99049 Armeria maritima Alba White Sea Thrift Armeria maritima Dusseldorf Pride Sea Thrift Armeria pseudarmeria Arrhenatherum elatius ssp. Bulbosum Variegatum Striped Tuber Oat Grass Artemisia Oriental Limelinght Wormwood Artemisia Powis Castle Wormwood Arum italicum Wild Ginger Aruncus aethusifolius Dwarf Korean Goat's Beard Arundo donax Variegata Striped Giant Reed Arundo donax Giant Reed Asparagus densiflorus Myers Asparagus Fern/Pony Tail Fern Asplenium scolopendrium Hart's Tounge Fern Aster dumosus Prof A. -

Chamaecyparis Obtusa (Hinoki Falsecypress) Hinoki Falsecypress Is a Conical-Shaped Evergreen Native to Japan
Chamaecyparis obtusa (Hinoki Falsecypress) Hinoki Falsecypress is a conical-shaped evergreen native to Japan. It has flat, fern-like, scaled leaves with white bands underneath. Its reddish-brown bark peels in long strips. There are many cultivars with different foliage coloration and growth habits. Landscape Information French Name: Cyprès du Japon, Hinoki Faux- Cyprès Pronounciation: kam-eh-SIP-uh-riss ob-TOO- suh Plant Type: Tree Origin: Japan Heat Zones: 1, 2, 3, 4, 5, 6, 7, 8 Hardiness Zones: 4, 5, 6, 7, 8 Uses: Screen, Hedge, Topiary, Bonsai, Espalier, Border Plant, Container, Windbreak Size/Shape Growth Rate: Slow Tree Shape: Pyramidal Canopy Symmetry: Symmetrical Canopy Density: Dense Canopy Texture: Fine Height at Maturity: Less than 0.5 m, 0.5 to 1 m, 1 to 1.5 m, 1.5 to 3 m, 3 to 5 m, 5 to 8 m, 8 to 15 m, 15 to 23 m Spread at Maturity: Less than 50 cm, 0.5 to 1 meter, 1 to 1.5 meters, 1.5 to 3 meters, 3 to 5 meters Plant Image Chamaecyparis obtusa (Hinoki Falsecypress) Botanical Description Foliage Leaf Arrangement: Opposite Leaf Venation: Nearly Invisible Leaf Persistance: Evergreen Leaf Type: Simple Leaf Blade: Less than 5 Leaf Shape: Scale Leaf Margins: Entire Leaf Textures: Medium Leaf Scent: Pleasant Color(growing season): Green Color(changing season): Green Leaf Image Flower Flower Showiness: False Flower Size Range: 0 - 1.5 Flower Scent: No Fragance Flower Color: Yellow Trunk Trunk Has Crownshaft: False Trunk Susceptibility to Breakage: Generally resists breakage Number of Trunks: Single Trunk Trunk Esthetic Values: Showy, -

Chamaecyparis Obtusa Hinoki Falsecypress1 Edward F
Fact Sheet ST-156 November 1993 Chamaecyparis obtusa Hinoki Falsecypress1 Edward F. Gilman and Dennis G. Watson2 INTRODUCTION This broad, sweeping, conical-shaped evergreen has graceful, flattened, fern-like branchlets which gently droop at branch tips (Fig. 1). Hinoki Falsecypress reaches 50 to 75 feet in height with a spread of 10 to 20 feet, has dark green foliage, and attractive, shredding, reddish-brown bark which peels off in long narrow strips. GENERAL INFORMATION Scientific name: Chamaecyparis obtusa Pronunciation: kam-eh-SIP-uh-riss ob-TOO-suh Common name(s): Hinoki Falsecypress Family: Cupressaceae USDA hardiness zones: 5 through 8A (Fig. 2) Origin: not native to North America Uses: Bonsai; screen Availability: somewhat available, may have to go out of the region to find the tree DESCRIPTION Height: 40 to 75 feet Spread: 10 to 20 feet Crown uniformity: symmetrical canopy with a regular (or smooth) outline, and individuals have more or less identical crown forms Figure 1. Mature Hinoki Falsecypress. Crown shape: pyramidal Crown density: dense Foliage Growth rate: medium Texture: fine Leaf arrangement: opposite/subopposite Leaf type: simple Leaf margin: entire 1. This document is adapted from Fact Sheet ST-156, a series of the Environmental Horticulture Department, Florida Cooperative Extension Service, Institute of Food and Agricultural Sciences, University of Florida. Publication date: November 1993. 2. Edward F. Gilman, associate professor, Environmental Horticulture Department; Dennis G. Watson, associate professor, Agricultural Engineering Department, Cooperative Extension Service, Institute of Food and Agricultural Sciences, University of Florida, Gainesville FL 32611. Chamaecyparis obtusa -- Hinoki Falsecypress Page 2 Figure 2. Shaded area represents potential planting range. -

Phylogeny of Hinterhubera, Novenia and Related
Louisiana State University LSU Digital Commons LSU Doctoral Dissertations Graduate School 2006 Phylogeny of Hinterhubera, Novenia and related genera based on the nuclear ribosomal (nr) DNA sequence data (Asteraceae: Astereae) Vesna Karaman Louisiana State University and Agricultural and Mechanical College, [email protected] Follow this and additional works at: https://digitalcommons.lsu.edu/gradschool_dissertations Recommended Citation Karaman, Vesna, "Phylogeny of Hinterhubera, Novenia and related genera based on the nuclear ribosomal (nr) DNA sequence data (Asteraceae: Astereae)" (2006). LSU Doctoral Dissertations. 2200. https://digitalcommons.lsu.edu/gradschool_dissertations/2200 This Dissertation is brought to you for free and open access by the Graduate School at LSU Digital Commons. It has been accepted for inclusion in LSU Doctoral Dissertations by an authorized graduate school editor of LSU Digital Commons. For more information, please [email protected]. PHYLOGENY OF HINTERHUBERA, NOVENIA AND RELATED GENERA BASED ON THE NUCLEAR RIBOSOMAL (nr) DNA SEQUENCE DATA (ASTERACEAE: ASTEREAE) A Dissertation Submitted to the Graduate Faculty of the Louisiana State University and Agricultural and Mechanical College in partial fulfillment of the requirements for the degree of Doctor of Philosophy in The Department of Biological Sciences by Vesna Karaman B.S., University of Kiril and Metodij, 1992 M.S., University of Belgrade, 1997 May 2006 "Treat the earth well: it was not given to you by your parents, it was loaned to you by your children. We do not inherit the Earth from our Ancestors, we borrow it from our Children." Ancient Indian Proverb ii ACKNOWLEDGMENTS I am indebted to many people who have contributed to the work of this dissertation. -

Rhinopithecus Brelichi): Patterns of Habitat Selection and Habitat Use
Global Ecology and Conservation 16 (2018) e00473 Contents lists available at ScienceDirect Global Ecology and Conservation journal homepage: http://www.elsevier.com/locate/gecco Original Research Article Altitudinal ranging of the Guizhou golden monkey (Rhinopithecus brelichi): Patterns of habitat selection and habitat use Yanqing Guo a, Jun Zhou a, 1, Jiahua Xie a, Paul A. Garber b, Michael Bruford c, * Baoping Ren d, Dayong Li e, Jiang Zhou a, a School of Life Sciences, Guizhou Normal University, Guiyang, Guizhou, China b Program in Ecology, Evolution, and Conservation Biology, University of Illinois, Urbana, IL, USA c Cardiff School of Biosciences and Sustainable Places Research Institute, Cadiff University, Cardiff, United Kingdom d Key Laboratory of Animal Ecology and Conservation Biology, Institute of Zoology, Chinese Academy of Sciences, Beijing, China e School of Life Sciences, China West Normal University, Nanchong, Sichuan, China article info abstract Article history: Between September 2011 and August 2013, we studied patterns of habitat selection and Received 17 August 2018 habitat use in the Guizhou golden monkey (Rhinopithecus brelichi) at Fanjingshan National Received in revised form 5 November 2018 Nature Reserve, China. the monkeys ranged across elevations between 1432 and 2100 m. Accepted 5 November 2018 Within this altitudinal range we recorded 125 genera, 72 families, and 236 tree and vine species. From these, the Guizhou golden monkey was observe to consume food items from Keywords: 104 species, 51 genera, and 31 plant families. Individual food species exploited by the Guizhou golden monkey monkeys varied significantly across seasons and by altitude. From October to March Altitudinal activity Habitat selection and utilization (Spring/Winter), the monkeys foraged across their entire 700 m elevation range. -

A Phylogenetic Analysis of Species Relationships in Hemlocks, the Genus Tsuga (Pinaceae)
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by East Tennessee State University East Tennessee State University Digital Commons @ East Tennessee State University Electronic Theses and Dissertations Student Works 8-2009 A Phylogenetic Analysis of Species Relationships in Hemlocks, the Genus Tsuga (Pinaceae). Jordan David Baker East Tennessee State University Follow this and additional works at: https://dc.etsu.edu/etd Part of the Paleobiology Commons Recommended Citation Baker, Jordan David, "A Phylogenetic Analysis of Species Relationships in Hemlocks, the Genus Tsuga (Pinaceae)." (2009). Electronic Theses and Dissertations. Paper 1791. https://dc.etsu.edu/etd/1791 This Thesis - Open Access is brought to you for free and open access by the Student Works at Digital Commons @ East Tennessee State University. It has been accepted for inclusion in Electronic Theses and Dissertations by an authorized administrator of Digital Commons @ East Tennessee State University. For more information, please contact [email protected]. A Phylogenetic Analysis of Species Relationships in Hemlocks, the Genus Tsuga (Pinaceae) __________________________ A thesis presented to the faculty of the Department of Biological Sciences East Tennessee State University In partial fulfillment of the requirements for the degree Master of Science in Biological Sciences _________________________ by Jordan Baker August 2009 ____________________ Timothy McDowell, Ph.D. - Chair Foster Levy, Ph.D. Christopher Liu, Ph.D. Keywords: Tsuga , Phylogeny, Morphology, Seedlings, Seed, Leaf Cuticle, Phylogenetic Analysis, Morphological Characters ABSTRACT A Phylogenetic Analysis of Species Relationships in Hemlocks, the Genus Tsuga (Pinaceae) by Jordan Baker The genus Tsuga is comprised of eight extant species found in North America and East Asia and four species represented by fossils from Europe and Japan. -

Astereae, Asteraceae) Downloaded from by Guest on 04 September 2019 GISELA SANCHO1*, PETER J
bs_bs_banner Botanical Journal of the Linnean Society, 2015, 177, 78–95. With 5 figures Late Cenozoic diversification of the austral genus Lagenophora (Astereae, Asteraceae) Downloaded from https://academic.oup.com/botlinnean/article-abstract/177/1/78/2416348 by guest on 04 September 2019 GISELA SANCHO1*, PETER J. DE LANGE FLS2, MARIANO DONATO3, JOHN BARKLA4 and STEVE J. WAGSTAFF5 1División Plantas Vasculares, Museo de La Plata, FCNYM, UNLP, Paseo del Bosque s.n., La Plata, 1900, Buenos Aires, Argentina 2Ecosystems and Species Unit, Department of Conservation, Private Bag 68908, Newton 1145, Auckland, New Zealand 3ILPLA, Instituto de Limnología Dr. Raúl A. Ringuelet, FCNYM, UNLP and CONICET, 122 and 60, La Plata, 1900, Buenos Aires, Argentina 4Otago Conservancy, Department of Conservation, PO Box 5244, Dunedin 9058, New Zealand 5Allan Herbarium, Landcare Research, PO Box 69040, Lincoln 7640, New Zealand Received 14 March 2014; revised 23 May 2014; accepted for publication 30 August 2014 Lagenophora (Astereae, Asteraceae) has 14 species in New Zealand, Australia, Asia, southern South America, Gough Island and Tristan da Cunha. Phylogenetic relationships in Lagenophora were inferred using nuclear and plastid DNA regions. Reconstruction of spatio-temporal evolution was estimated using parsimony, Bayesian inference and likelihood methods, a Bayesian relaxed molecular clock and ancestral area and habitat reconstruc- tions. Our results support a narrow taxonomic concept of Lagenophora including only a core group of species with one clade diversifying in New Zealand and another in South America. The split between the New Zealand and South American Lagenophora dates from 11.2 Mya [6.1–17.4 95% highest posterior density (HPD)].