Recurrent Bridgehead Effects Accelerate Global Alien Ant Spread
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Winged Ants (Hymenoptera: Formicidae) Presence in Twigs on the Leaf Litter of Atlantic Forest
Biota Neotropica 19(3): e20180694, 2019 www.scielo.br/bn ISSN 1676-0611 (online edition) Inventory Winged ants (Hymenoptera: Formicidae) presence in twigs on the leaf litter of Atlantic Forest Tae Tanaami Fernandes1 , Rogério R. Silva2, Débora Rodrigues de Souza-Campana2, Otávio Guilherme Morais da Silva2 & Maria Santina de Castro Morini1* 1Universidade de Mogi das Cruzes, Laboratório de Mirmecologia do Alto Tietê, Rua Dr. Cândido Xavier de Almeida e Souza, 200, CEP 08780-911, Mogi das Cruzes, SP, Brasil 2Museu Paraense Emílio Goeldi, Coordenação de Ciências da Terra e Ecologia, Avenida Perimetral, 1901, Terra Firme, CEP 66077-830, Belém, PA, Brasil *Corresponding author: Maria Santina de Castro Morini, e-mail: [email protected] FERNANDES, T. T., SILVA, R. R., SOUZA-CAMPANA, D. R., SILVA, O. G. M., MORINI, M. S. C. Winged ants (Hymenoptera: Formicidae) presence in twigs on the leaf litter of Atlantic Forest. Biota Neotropica 19(3): e20180694. http://dx.doi.org/10.1590/1676-0611-BN-2018-0694 Abstract: In the leaf litter, ants have various nesting resources available, such as live or dead trunks, twigs, leaves, fruits and seeds. On the twigs, there are adults and immature individuals, but also the queen and winged. The production of wings requires time and energy from the colony. The objective of this study was to investigate the presence of winged in ant colonies in twigs on the leaf litter. Our prediction is that the richness and abundance of winged in twigs are the greatest in rainy months. We collected all twigs with ants in 552 plots with 16 m2, totaling 8,832 m2 of leaf litter, in areas located in the Brazilian Atlantic Domain. -

ORIGINAL ARTICLE a Novel Intracellular Mutualistic Bacterium in the Invasive Ant Cardiocondyla Obscurior
The ISME Journal (2016) 10, 376–388 © 2016 International Society for Microbial Ecology All rights reserved 1751-7362/16 www.nature.com/ismej ORIGINAL ARTICLE A novel intracellular mutualistic bacterium in the invasive ant Cardiocondyla obscurior Antonia Klein1,8, Lukas Schrader1,8, Rosario Gil2, Alejandro Manzano-Marín2, Laura Flórez3, David Wheeler5, John H Werren6, Amparo Latorre2,7, Jürgen Heinze1, Martin Kaltenpoth3,4, Andrés Moya2 and Jan Oettler1 1Institut für Zoologie, Universität Regensburg, Regensburg, Germany; 2Institut Canvanilles de Biodiversitat i Biologia Evolutiva (ICBiBE), Parc Cientific de la Universitat de Valencia, Paterna (Valencia), Spain; 3Max Planck Institute for Chemical Ecology, Jena, Germany; 4Johannes Gutenberg University Mainz, Institute for Zoology, Department for Evolutionary Ecology, Mainz, Germany; 5Institute of Fundamental Sciences, Massey University, Palmerston North, New Zealand; 6Department of Biology, University Rochester, Rochester, NY, USA and 7Área de Genómica y Salud de la Fundación para el Fomento de la Investigación Sanitaria y Biomédica de la Comunitat Valenciana (FISABIO)–Salud Pública, Valencia, Spain The evolution of eukaryotic organisms is often strongly influenced by microbial symbionts that confer novel traits to their hosts. Here we describe the intracellular Enterobacteriaceae symbiont of theinvasiveantCardiocondyla obscurior, ‘Candidatus Westeberhardia cardiocondylae’. Upon metamorphosis, Westeberhardia is found in gut-associated bacteriomes that deteriorate following eclosion. Only queens maintain Westeberhardia in the ovarian nurse cells from where the symbionts are transmitted to late-stage oocytes during nurse cell depletion. Functional analyses of the streamlined genome of Westeberhardia (533 kb, 23.41% GC content) indicate that neither vitamins nor essential amino acids are provided for the host. However, the genome encodes for an almost complete shikimate pathway leading to 4-hydroxyphenylpyruvate, which could be converted into tyrosine by the host. -

Taxonomy and Distribution of the Argentine Ant, Linepithema Humile (Hymenoptera: Formicidae)
SYSTEMATICS Taxonomy and Distribution of the Argentine Ant, Linepithema humile (Hymenoptera: Formicidae) ALEXANDER L. WILD Department of Entomology, University of California at Davis, Davis, CA 95616 Ann. Entomol. Soc. Am. 97(6): 1204Ð1215 (2004) ABSTRACT The taxonomy of an invasive pest species, the Argentine ant, is reviewed. Linepithema humile (Mayr) 1868 is conÞrmed as the valid name for the Argentine ant. Morphological variation and species boundaries of L. humile are examined, with emphasis on populations from the antÕs native range in southern South America. Diagnoses and illustrations are provided for male, queen, and worker castes. Collection records of L. humile in South America support the idea of a native distribution closely associated with major waterways in lowland areas of the Parana´ River drainage, with recent intro- ductions into parts of Argentina, Brazil, Chile, Colombia, Ecuador, and Peru. KEY WORDS Argentine ant, Linepithema humile, taxonomy, invasive species THE ARGENTINE ANT, Linepithema humile (Mayr) 1868, MCSN, MCZC, MHNG, MZSP, NHMB, NHMW, and is among the worldÕs most successful invasive species. USNM; see below for explanation of abbreviations). This native South American insect has become a cos- Taxonomic confusion over L. humile extends be- mopolitan pest, particularly in the Mediterranean cli- yond museum collections. At least one important mates of North America, Chile, South Africa, Austra- study, seeking to explain Argentine ant population lia, and southern Europe (Suarez et al. 2001). regulation in the native range through phorid para- Argentine ants have been implicated in the decline of sitism (Orr and Seike 1998), initially targeted the native arthropod (Cole et al. 1992) and vertebrate wrong Linepithema species (Orr et al. -

James K. Wetterer
James K. Wetterer Wilkes Honors College, Florida Atlantic University 5353 Parkside Drive, Jupiter, FL 33458 Phone: (561) 799-8648; FAX: (561) 799-8602; e-mail: [email protected] EDUCATION UNIVERSITY OF WASHINGTON, Seattle, WA, 9/83 - 8/88 Ph.D., Zoology: Ecology and Evolution; Advisor: Gordon H. Orians. MICHIGAN STATE UNIVERSITY, East Lansing, MI, 9/81 - 9/83 M.S., Zoology: Ecology; Advisors: Earl E. Werner and Donald J. Hall. CORNELL UNIVERSITY, Ithaca, NY, 9/76 - 5/79 A.B., Biology: Ecology and Systematics. UNIVERSITÉ DE PARIS III, France, 1/78 - 5/78 Semester abroad: courses in theater, literature, and history of art. WORK EXPERIENCE FLORIDA ATLANTIC UNIVERSITY, Wilkes Honors College 8/04 - present: Professor 7/98 - 7/04: Associate Professor Teaching: Biodiversity, Principles of Ecology, Behavioral Ecology, Human Ecology, Environmental Studies, Tropical Ecology, Field Biology, Life Science, and Scientific Writing 9/03 - 1/04 & 5/04 - 8/04: Fulbright Scholar; Ants of Trinidad and Tobago COLUMBIA UNIVERSITY, Department of Earth and Environmental Science 7/96 - 6/98: Assistant Professor Teaching: Community Ecology, Behavioral Ecology, and Tropical Ecology WHEATON COLLEGE, Department of Biology 8/94 - 6/96: Visiting Assistant Professor Teaching: General Ecology and Introductory Biology HARVARD UNIVERSITY, Museum of Comparative Zoology 8/91- 6/94: Post-doctoral Fellow; Behavior, ecology, and evolution of fungus-growing ants Advisors: Edward O. Wilson, Naomi Pierce, and Richard Lewontin 9/95 - 1/96: Teaching: Ethology PRINCETON UNIVERSITY, Department of Ecology and Evolutionary Biology 7/89 - 7/91: Research Associate; Ecology and evolution of leaf-cutting ants Advisor: Stephen Hubbell 1/91 - 5/91: Teaching: Tropical Ecology, Introduction to the Scientific Method VANDERBILT UNIVERSITY, Department of Psychology 9/88 - 7/89: Post-doctoral Fellow; Visual psychophysics of fish and horseshoe crabs Advisor: Maureen K. -

Population Dynamics of Invasive Argentine Ant
BioInvasions Records (2021) Volume 10, Issue 2: 467–476 CORRECTED PROOF Research Article Population dynamics of invasive Argentine ant Linepithema humile Mayr, 1868 (Hymenoptera: Formicidae) haplotypes in Kobe Port, Japan, and implications for the prediction of future dispersal and effective management Yugo Seko1,#,*, Yu Maebara2,#, Naoyuki Nakahama3,4, Takuya Nakamori1, Naoto Ishiwaka5, Yuki Morikawa5, Daisuke Hayasaka1,* and Takuo Sawahata1 1Graduate school of Agriculture, Kindai University, 3327-204, Nakamachi, Nara City, Nara Prefecture, Japan 2Nagoya Branch Office, Nippon Koei Co. Ltd., 1-20-22, Aoi, Naka-ku, Nagoya City, Aichi Prefecture, Japan 3Institute of Natural and Environmental Sciences, University of Hyogo, 6 Yayoigaoka, Sanda City, Hyogo Prefecture, Japan 4Division of Ecological Restoration, The Museum of Nature and Human Activities, 6 Yayoigaoka, Sanda City, Hyogo Prefecture, Japan 5Faculty of Agriculture, Kindai University, 3327-204, Nakamachi, Nara City, Nara Prefecture, Japan Author e-mails: [email protected] (YS), [email protected] (YMa), [email protected] (NN), [email protected] (TN), [email protected] (NI), [email protected] (YMo), [email protected]; [email protected] (DH), [email protected] (TS) #These authors contributed equally *Corresponding authors Citation: Seko Y, Maebara Y, Nakahama N, Nakamori T, Ishiwaka N, Morikawa Y, Abstract Hayasaka D, Sawahata T (2021) Population dynamics of invasive Argentine The Argentine ant Linepithema humile Mayr, native to South America, has been ant Linepithema humile Mayr, 1868 unintentionally introduced worldwide. Although L. humile usually forms an extremely (Hymenoptera: Formicidae) haplotypes in large supercolony all sharing the same haplotype among members in its introduced Kobe Port, Japan, and implications for the ranges, four haplotypes (LH1, LH2, LH3, LH4) with different genetic structures prediction of future dispersal and effective management. -

Encyclopedia of Social Insects
G Guests of Social Insects resources and homeostatic conditions. At the same time, successful adaptation to the inner envi- Thomas Parmentier ronment shields them from many predators that Terrestrial Ecology Unit (TEREC), Department of cannot penetrate this hostile space. Social insect Biology, Ghent University, Ghent, Belgium associates are generally known as their guests Laboratory of Socioecology and Socioevolution, or inquilines (Lat. inquilinus: tenant, lodger). KU Leuven, Leuven, Belgium Most such guests live permanently in the host’s Research Unit of Environmental and nest, while some also spend a part of their life Evolutionary Biology, Namur Institute of cycle outside of it. Guests are typically arthropods Complex Systems, and Institute of Life, Earth, associated with one of the four groups of eusocial and the Environment, University of Namur, insects. They are referred to as myrmecophiles Namur, Belgium or ant guests, termitophiles, melittophiles or bee guests, and sphecophiles or wasp guests. The term “myrmecophile” can also be used in a broad sense Synonyms to characterize any organism that depends on ants, including some bacteria, fungi, plants, aphids, Inquilines; Myrmecophiles; Nest parasites; and even birds. It is used here in the narrow Symbionts; Termitophiles sense of arthropods that associated closely with ant nests. Social insect nests may also be parasit- Social insect nests provide a rich microhabitat, ized by other social insects, commonly known as often lavishly endowed with long-lasting social parasites. Although some strategies (mainly resources, such as brood, retrieved or cultivated chemical deception) are similar, the guests of food, and nutrient-rich refuse. Moreover, nest social insects and social parasites greatly differ temperature and humidity are often strictly regu- in terms of their biology, host interaction, host lated. -

Impact of the Invasive Crazy Ant Anoplolepis Gracilipes on Bird Island, Seychelles
Journal of Insect Conservation 8: 15–25, 2004. 15 # 2004 Kluwer Academic Publishers. Printed in the Netherlands. Impact of the invasive crazy ant Anoplolepis gracilipes on Bird Island, Seychelles J. Gerlach University Museum of Zoology Cambridge, Department of Zoology, Downing Street, Cambridge CB2 3EJ, UK (e-mail: [email protected]) Received 26 March 2003; accepted in revised form 12 December 2003 Key words: Anoplolepis gracilipes, Ant, Formicidae, Invasion, Management, Seychelles Abstract The crazy ant (Anoplolepis gracilipes) invaded Bird island, Seychelles, in the 1980s. In 1997, its range expanded and population densities increased. The impacts of this change were studied in 2001 using a combination of arthropod collecting methods. The ant population excluded larger invertebrates (principally the large ant Odontomachus simillimus and the crabs, principally Ocypode spp.). Cockroaches, however, remained abundant in ant-infested areas and tree-nesting birds (Lesser Noddy Anous tenuirostris) appear to be able to breed successfully in the presence of the crazy ant. The ants are only abundant in areas of deep shade which provide cool nesting areas, yet enabling them to forage in the open when ground temperatures fall. The expansion of the ants was correlated with the regeneration of woodland on the island. Recommendations are made for the management of the woodland which may reduce the impacts of the crazy ant. Introduction Africa, Asia, America and Australia (Dorow 1996). This species has been recorded as excluding The threat from alien invasive species is widely other ant species (Greenslade 1971) and recently perceived as one of the major contributions to has been implicated in a population crash in the present-day high-extinction rates, affecting an esti- red crab Gecarcoidea natalis on Christmas Island mated 30% of threatened birds, 15% of threatened (O’ Dowd et al. -

Genetic Mechanisms Underlying the Evolutionary Success of Eusocial Insects
insects Review (Epi)Genetic Mechanisms Underlying the Evolutionary Success of Eusocial Insects Kayli R. Sieber 1 , Taylor Dorman 1, Nicholas Newell 1 and Hua Yan 1,2,* 1 Department of Biology, University of Florida, Gainesville, FL 32611, USA; kayli.sieber@ufl.edu (K.R.S.); taylor.dorman@ufl.edu (T.D.); nicholas.newell@ufl.edu (N.N.) 2 Center for Smell and Taste, University of Florida, Gainesville, FL 32611, USA * Correspondence: hua.yan@ufl.edu; Tel.: +1-352-273-4983 Simple Summary: Social insects, namely ants, bees, and termites, are among the most numerous and successful animals on Earth. This is due to a variety of features: highly cooperative behavior performed by colony members and their specialization on a variety of tasks. Diverse physiological and behavioral specializations are regulated not only by the genetic system, but also by the epige- netic system which alters gene expressions without modifying the genetic code. This review will summarize recent advancements in such studies in eusocial insects. Abstract: Eusocial insects, such as bees, ants, and wasps of the Hymenoptera and termites of the Blattodea, are able to generate remarkable diversity in morphology and behavior despite being genetically uniform within a colony. Most eusocial insect species display caste structures in which reproductive ability is possessed by a single or a few queens while all other colony members act Citation: Sieber, K.R.; Dorman, T.; as workers. However, in some species, caste structure is somewhat plastic, and individuals may Newell, N.; Yan, H. (Epi)Genetic switch from one caste or behavioral phenotype to another in response to certain environmental cues. -
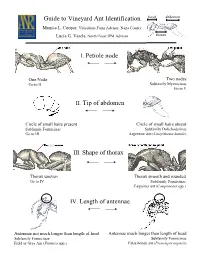
I. Petiole Node II. Tip of Abdomen IV. Length of Antennae Guide to Vineyard Ant Identification III. Shape of Thorax
Guide to Vineyard Ant Identification head abdomen Monica L. Cooper, Viticulture Farm Advisor, Napa County Lucia G. Varela, North Coast IPM Advisor thorax I. Petiole node One Node Two nodes Go to II Subfamily Myrmicinae Go to V II. Tip of abdomen Circle of small hairs present Circle of small hairs absent Subfamily Formicinae Subfamily Dolichoderinae Go to III Argentine Ant (Linepithema humile) III. Shape of thorax Thorax uneven Thorax smooth and rounded Go to IV Subfamily Formicinae Carpenter ant (Camponotus spp.) IV. Length of antennae Antennae not much longer than length of head Antennae much longer than length of head Subfamily Formicinae Subfamily Formicinae Field or Gray Ant (Formica spp.) False honey ant (Prenolepis imparis) head abdomen Petiole with two nodes Subfamily Myrmicinae (V-VIII) thorax V. Dorsal side of Thorax & Antennae One pair of spines on thorax No spines on thorax 12 segmented antennae 10 segmented antennae Go to VI Solenopsis molesta and Solenopsis xyloni VI. Underside of head No brush of bristles Brush of long bristles Go to VII Harvester ants (Pogonomyrmex californicus and P. brevispinosis) VII. Head and Thorax With hairs Without hairs Go to VIII Cardiocondyla mauritanica VIII. Head and Thorax With many parallel furrows Without parallel furrows Profile of thorax rounded Profile of thorax not evenly rounded Pavement ant (Tetramorium “species E”) Pheidole californica Argentine Ant (Linepithema humile), subfamily Dolichoderinae Exotic species 3-4 mm in length Deep brown to light black Move rapidly in distinct trails Feed on honeydew Shallow nests (2 inches from soil surface) Alex Wild Does not bite or sting Carpenter Ant (Camponotus spp.), subfamily Formicinae Large ant: >6 mm in length Dark color with smooth, rounded thorax Workers most active at dusk and night One of most abundant and widespread genera worldwide Generalist scavengers and predators: feed on dead and living insects, nectar, fruit juices and Jack K. -

Hymenoptera: Formicidae: Dolichoderinae) from Colombia
Revista Colombiana de Entomología 37 (1): 159-161 (2011) 159 Scientific note The first record of the genus Gracilidris (Hymenoptera: Formicidae: Dolichoderinae) from Colombia Primer registro del género Gracilidris (Hymenoptera: Formicidae: Dolichoderinae) para Colombia ROBERTO J. GUERRERO1 and CATALINA SANABRIA2 Abstract: The dolichoderine ant genus Gracilidris and its sole species, G. pombero, are recorded for the first time for Colombia from populations from the foothills of the Colombian Amazon basin. Comments and hypotheses about the biogeography of the genus are discussed. Key words: Ants. Biodiversity. Caquetá. Colombian Amazon. Grazing systems. Resumen: El género dolicoderino de hormigas Gracilidris y su única especie, G. pombero, son registrados por primera vez para Colombia, de poblaciones provenientes del piedemonte de la cuenca Amazónica colombiana. Algunos comen- tarios e hipótesis sobre la biogeografía del género son discutidos. Palabras clave: Hormigas. Biodiversidad. Caquetá. Amazonas colombiano. Pasturas ganaderas. Introduction distribution of the genus Gracilidris in South America. We also discuss each of the dolichoderine genera that occur in Currently, dolichoderine ants (Hymenoptera: Formicidae: Colombia. Dolichoderinae) include 28 extant genera (Bolton et al. 2006; Fisher 2009) distributed in four tribes according to Methods the latest higher level classification of the ant subfamily Dolichoderinae (Ward et al. 2010). Eleven of those extant We separated G. pombero specimens from all ants collected genera occur in the New World: Bothriomyrmex, Dolicho- in the project “Amaz_BD: Biodiversidad de los paisajes derus, Liometopum, Tapinoma, and Technomyrmex have a Amazónicos, determinantes socio-económicos y produc- global distribution, while Anillidris, Azteca, Dorymyrmex, ción de bienes y servicios”. This research was carried out in Forelius, Gracilidris, and Linepithema are endemic to the Caquetá department located in the western foothills of the New World. -

A Pugnacious Ant (Anoplolepis Custodiens)
A Pugnacious ant (Anoplolepis custodiens) confounds ant assemblage responses to bush encroachment Thinandavha Munyai1, Stefan Foord2, Rob Slotow1, and Nomathamsanqa Mkhize1 1University of KwaZulu-Natal College of Agriculture Engineering and Science 2University of Venda November 25, 2020 Abstract Habitat structure is a key determinant of variation in biodiversity. The effects of increased vertical and horizontal vegetation structure can result in marked shifts in animal communities. This is particularly true for ants in response to woody thickening, with predicted negative impacts on ant diversity. We used pitfall traps to study the response of epigeic ants in two co-occurring dominant habitats (closed and open) of an African savanna biome experiencing extensive woody thickening. Although species richness was higher in open habitats, evenness was significantly lower. Thickening explained significant amounts of variation in ant composition, but site-specific characteristics and seasonality were more important. These site-specific characteristics were largely linked to Anoplolepis custodiens, a species that were locally abundant in open habitats with clayey soils, where they often accounted for more than 90% of all ant activity. As A. custodiens also responds positively to disturbance, indiscriminate bush clearing could lead to knock on effects associated with the numerical and behavioural dominance of this species. Introduction The savanna biome dominates southern Africa, and is characterised by continuous grassy understory, a discontinuous woody overstory (Archer et al., 2017; Bond, 2019), and, therefore, considered a largely open ecosystem (Pausas & Bond, 2020). However, various anthropogenic activities (Osborne et al., 2018) pose a threat to this biome. One pervasive threat is that of woody plant encroachment into savannas and grasslands, linked to climate change in particular (Criado, Myers-Smith, Bjorkman, Lehmann, & Stevens, 2020). -

Use of an Argentine Ant, Linepithema Humile, Semiochemical to Deliver an Acute Toxicant
insects Article Use of an Argentine Ant, Linepithema humile, Semiochemical to Deliver an Acute Toxicant Benjamin M. Gochnour 1, Daniel R. Suiter 2,*, Jerry W. Davis 3 and Qingguo Huang 4 1 Department of Entomology, University of Georgia, 120 Cedar Street, Athens, GA 30602, USA; [email protected] 2 Department of Entomology, University of Georgia Griffin Camus, 1109 Experiment Street, Griffin, GA 30223, USA 3 Experimental Statistics, College of Agricultural and Environmental Sciences, University of Georgia, 1109 Experiment Street, Griffin, GA 30223, USA; [email protected] 4 Department of Crop and Soil Sciences, University of Georgia Griffin Campus, 1109 Experiment Street, Griffin, GA 30223, USA; [email protected] * Correspondence: [email protected]; Tel.: +1-770-233-6114 Received: 14 October 2018; Accepted: 16 November 2018; Published: 23 November 2018 Abstract: The Argentine ant, Linepithema humile, is an invasive nuisance, agricultural, and ecological pest from South America. In the United States, its primary distribution is in California and the Southeast. The structural pest control industry responds to property owner complaints when this ant’s populations become problematic and a persistent nuisance. Actions taken to control Argentine ants in the urban and suburban environment are typically complaint-driven, and often involve the application of insecticide sprays applied to the outdoor environment by professional pest managers. In California, and elsewhere, spray treatments of various residual insecticides by property owners and pest management professionals has resulted in significant runoff and in subsequent surface water contamination. As a result, an immediate need exists to develop alternative methods of ant control targeted at reducing environmental contamination.