Historical and Current Forest and Range Landscapes in the Interior
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Forests of Eastern Oregon: an Overview Sally Campbell, Dave Azuma, and Dale Weyermann
Forests of Eastern Oregon: An Overview Sally Campbell, Dave Azuma, and Dale Weyermann United States Forest Pacific Northwest General Tecnical Report Department of Service Research Station PNW-GTR-578 Agriculture April 2003 Revised 2004 Joseph area, eastern Oregon. Photo by Tom Iraci Authors Sally Campbell is a biological scientist, Dave Azuma is a research forester, and Dale Weyermann is geographic information system manager, U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station, 620 SW Main, Portland, OR 97205. Cover: Aspen, Umatilla National Forest. Photo by Tom Iraci Forests of Eastern Oregon: An Overview Sally Campbell, Dave Azuma, and Dale Weyermann U.S. Department of Agriculture Forest Service Pacific Northwest Research Station Portland, OR April 2003 State Forester’s Welcome Dear Reader: The Oregon Department of Forestry and the USDA Forest Service invite you to read this overview of eastern Oregon forests, which provides highlights from recent forest inventories.This publication has been made possible by the USDA Forest Service Forest Inventory and Analysis (FIA) Program, with support from the Oregon Department of Forestry. This report was developed from data gathered by the FIA in eastern Oregon’s forests in 1998 and 1999, and has been supplemented by inventories from Oregon’s national forests between 1993 and 1996.This report and other analyses of FIA inventory data will be extremely useful as we evaluate fire management strategies, opportunities for improving rural economies, and other elements of forest management in eastern Oregon.We greatly appreciate FIA’s willingness to work with the researchers, analysts, policymakers, and the general public to collect, analyze, and distrib- ute information about Oregon’s forests. -

Flood Basalts and Glacier Floods—Roadside Geology
u 0 by Robert J. Carson and Kevin R. Pogue WASHINGTON DIVISION OF GEOLOGY AND EARTH RESOURCES Information Circular 90 January 1996 WASHINGTON STATE DEPARTMENTOF Natural Resources Jennifer M. Belcher - Commissioner of Public Lands Kaleen Cottingham - Supervisor FLOOD BASALTS AND GLACIER FLOODS: Roadside Geology of Parts of Walla Walla, Franklin, and Columbia Counties, Washington by Robert J. Carson and Kevin R. Pogue WASHINGTON DIVISION OF GEOLOGY AND EARTH RESOURCES Information Circular 90 January 1996 Kaleen Cottingham - Supervisor Division of Geology and Earth Resources WASHINGTON DEPARTMENT OF NATURAL RESOURCES Jennifer M. Belcher-Commissio11er of Public Lands Kaleeo Cottingham-Supervisor DMSION OF GEOLOGY AND EARTH RESOURCES Raymond Lasmanis-State Geologist J. Eric Schuster-Assistant State Geologist William S. Lingley, Jr.-Assistant State Geologist This report is available from: Publications Washington Department of Natural Resources Division of Geology and Earth Resources P.O. Box 47007 Olympia, WA 98504-7007 Price $ 3.24 Tax (WA residents only) ~ Total $ 3.50 Mail orders must be prepaid: please add $1.00 to each order for postage and handling. Make checks payable to the Department of Natural Resources. Front Cover: Palouse Falls (56 m high) in the canyon of the Palouse River. Printed oo recycled paper Printed io the United States of America Contents 1 General geology of southeastern Washington 1 Magnetic polarity 2 Geologic time 2 Columbia River Basalt Group 2 Tectonic features 5 Quaternary sedimentation 6 Road log 7 Further reading 7 Acknowledgments 8 Part 1 - Walla Walla to Palouse Falls (69.0 miles) 21 Part 2 - Palouse Falls to Lower Monumental Dam (27.0 miles) 26 Part 3 - Lower Monumental Dam to Ice Harbor Dam (38.7 miles) 33 Part 4 - Ice Harbor Dam to Wallula Gap (26.7 mi les) 38 Part 5 - Wallula Gap to Walla Walla (42.0 miles) 44 References cited ILLUSTRATIONS I Figure 1. -

The Harney County Way Collaborative Summit May 2 – 3, 2018 | Lincoln Building Auditorium, Burns, OR
The Harney County Way Collaborative Summit May 2 – 3, 2018 | Lincoln Building Auditorium, Burns, OR The Harney County Way Collaborative Summit 2018 Linking Collaboration Efforts to Build a Best Harney County High Desert Partnership’s Mission Summit Vision The High Desert Partnership exists to cultivate collaboration and We believe this summit will provide a productive time for those support and strengthen diverse partners engaged in solving participating in collaborative work in Harney County to network, complex issues to advance healthy ecosystems, economic well- learn and look for opportunities to work together. Bringing together being and social vitality to ensure a thriving and resilient the collaborative initiatives will create synergy and the story of community. collaboration will reverberate in our community. The outcomes from this summit will lead to more resilient communities. Our Core Values o We believe in our collaborative process to address societal Goals of the Summit issues. o Increase the understanding of collaborative efforts in Harney o We believe in doing things right rather than right now. County. o We believe in recognizing the values of others. o Understand the links where initiatives can work together on o We believe in advocating for the process, not for outcomes. projects or programs. o We believe in taking a holistic approach: social, ecological and o Find the places for sharing resources. economic. o Grow the community's collaborative participation. o We believe that optimism is necessary to successfully address o Provide a venue for those in attendance to gain a better the challenges we face. understanding of the work of High Desert Partnership. -

Characterization of Ecoregions of Idaho
1 0 . C o l u m b i a P l a t e a u 1 3 . C e n t r a l B a s i n a n d R a n g e Ecoregion 10 is an arid grassland and sagebrush steppe that is surrounded by moister, predominantly forested, mountainous ecoregions. It is Ecoregion 13 is internally-drained and composed of north-trending, fault-block ranges and intervening, drier basins. It is vast and includes parts underlain by thick basalt. In the east, where precipitation is greater, deep loess soils have been extensively cultivated for wheat. of Nevada, Utah, California, and Idaho. In Idaho, sagebrush grassland, saltbush–greasewood, mountain brush, and woodland occur; forests are absent unlike in the cooler, wetter, more rugged Ecoregion 19. Grazing is widespread. Cropland is less common than in Ecoregions 12 and 80. Ecoregions of Idaho The unforested hills and plateaus of the Dissected Loess Uplands ecoregion are cut by the canyons of Ecoregion 10l and are disjunct. 10f Pure grasslands dominate lower elevations. Mountain brush grows on higher, moister sites. Grazing and farming have eliminated The arid Shadscale-Dominated Saline Basins ecoregion is nearly flat, internally-drained, and has light-colored alkaline soils that are Ecoregions denote areas of general similarity in ecosystems and in the type, quality, and America into 15 ecological regions. Level II divides the continent into 52 regions Literature Cited: much of the original plant cover. Nevertheless, Ecoregion 10f is not as suited to farming as Ecoregions 10h and 10j because it has thinner soils. -

Final Environmental Impact Statement and Proposed Land-Use Plan Amendments for the Boardman to Hemingway Transmission Line Proje
B2H Final EIS and Proposed LUP Amendments Appendix H—Visual Resources Supporting Data Appendix H VISUAL RESOURCES SUPPORTING DATA This appendix includes the following: Appendix H1 – Visual Analysis Unit Descriptions - Visual Analysis Unit Descriptions Table - Change in Cultural Modification to the Scenic Quality Rating Units Appendix H2 – Contrast Rating Worksheets - Baker Field Office Visual Contrast Rating Worksheets* - Malheur Field Office Visual Contrast Rating Worksheets* - Owyhee Field Office Visual Contrast Rating Worksheets* - Additional Visual Contrast Rating Worksheets Appendix H3 – Photo Simulations - Photo Simulations from Visual Resource Report 1 - Additional Photo Simulations *NOTE: For the Final Environmental Impact Statement, additional route variations have been analyzed. As a result, certain routes analyzed for the Draft Environmental Impact Statement have been renamed. They are as follows: Proposed Action changed to Applicant’s Proposed Action Alternative Burnt River Alternative to Flagstaff A – Burnt River Alternative Flagstaff Hill to Flagstaff A Alternative Double Mountain Alternative to Variation S5-B2 H-1 This page intentionally left blank. B2H Final EIS and Proposed LUP Amendments Appendix H1—Visual Analysis Unit Descriptions Appendix H1 VISUAL ANALYSIS UNIT DESCRIPTIONS The Visual Analysis Unit (VAU) descriptions are provided in Table H1-1, which includes an overall description of each VAU within the B2H Project area for visual resources. The descriptions of the units include information about the landforms, topography, water, and vegetation within the units, as well as other features and information. The VAUs are identified by two digits, followed by three numbers, and a unit name. The two digits represent the BLM field office or resource area in which the unit is located (BR=Border Resource Area; CE=Central Oregon Resource Area; BA=Baker Resource Area; MA=Malheur Resource Area; OW=Owyhee Field Office; FR=Four Rivers Field Office). -

Summary of the Columbia Plateau Regional Aquifer-System Analysis, Washington, Oregon, and Idaho
SUMMARY OF THE COLUMBIA PLATEAU REGIONAL AQUIFER-SYSTEM ANALYSIS, WASHINGTON, OREGON, AND IDAHO PROFESSIONAL PAPER 1413-A USGS tience for a changing world Availability of Publications of the U.S. Geological Survey Order U.S. Geological Survey (USGS) publications from the Documents. Check or money order must be payable to the offices listed below. Detailed ordering instructions, along with Superintendent of Documents. Order by mail from prices of the last offerings, are given in the current-year issues of the catalog "New Publications of the U.S. Geological Superintendent of Documents Survey." Government Printing Office Washington, DC 20402 Books, Maps, and Other Publications Information Periodicals By Mail Many Information Periodicals products are available through Books, maps, and other publications are available by mail the systems or formats listed below: from Printed Products USGS Information Services Box 25286, Federal Center Printed copies of the Minerals Yearbook and the Mineral Com Denver, CO 80225 modity Summaries can be ordered from the Superintendent of Publications include Professional Papers, Bulletins, Water- Documents, Government Printing Office (address above). Supply Papers, Techniques of Water-Resources Investigations, Printed copies of Metal Industry Indicators and Mineral Indus Circulars, Fact Sheets, publications of general interest, single try Surveys can be ordered from the Center for Disease Control copies of permanent USGS catalogs, and topographic and and Prevention, National Institute for Occupational Safety and thematic maps. Health, Pittsburgh Research Center, P.O. Box 18070, Pitts burgh, PA 15236-0070. Over the Counter Mines FaxBack: Return fax service Books, maps, and other publications of the U.S. Geological Survey are available over the counter at the following USGS 1. -
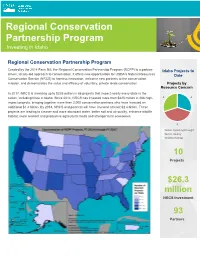
Regional Conservation Partnership Program Investing in Idaho
Regional Conservation Partnership Program Investing in Idaho Regional Conservation Partnership Program Created by the 2014 Farm Bill, the Regional Conservation Partnership Program (RCPP) is a partner- Idaho Projects to driven, locally-led approach to conservation. It offers new opportunities for USDA’s Natural Resources Date Conservation Service (NRCS) to harness innovation, welcome new partners to the conservation mission, and demonstrates the value and efficacy of voluntary, private lands conservation. Projects by Resource Concern In 2017, NRCS is investing up to $225 million in 88 projects that impact nearly every state in the nation, including three in Idaho. Since 2014, NRCS has invested more than $825 million in 286 high- 3 3 impact projects, bringing together more than 2,000 conservation partners who have invested an additional $1.4 billion. By 2018, NRCS and partners will have invested at least $2.4 billion. These projects are leading to cleaner and more abundant water, better soil and air quality, enhance wildlife habitat, more resilient and productive agricultural lands and stronger rural economies. 4 Water Quantity/Drought Water Quality Wildlife Habitat 10 Projects $26.3 million NRCS Investment 93 Partners Idaho RCPP Projects Existing RCPP Projects Year Title Funding Lead Partner Number of NRCS Pool Partners Investment 2016 Farmer's Cooperative Ditch Company Project State Farmer’s Cooperative Ditch 7 $500,000 Company 2016 Greater Spokane River Watershed National Spokane Conservation 21 $7.7 million Implementation District 2016 -

Iron Horse State Park Master Plan Addendum Iron Horse Palouse – Malden to Idaho Border
MASTER PLAN IRON HORSE STATE PARK MASTER PLAN ADDENDUM IRON HORSE PALOUSE – MALDEN TO IDAHO BORDER WASHINGTON STATE PARKS AND RECREATION COMMISSION Approved March 27, 2014 TABLE OF CONTENTS CHAPTER 1: INTRODUCTION ......................................................................................................................... 1 PURPOSE ................................................................................................................................................... 1 STUDY AREA .............................................................................................................................................. 1 RECREATION USE ...................................................................................................................................... 3 RECREATION SERVICE AREA ...................................................................................................................... 3 CHAPTER 2: EXISTING CONDITIONS .............................................................................................................. 5 STUDY APPROACH ..................................................................................................................................... 5 LAND OWNERSHIP .................................................................................................................................... 5 EXISTING BRIDGES ..................................................................................................................................... 5 REMOVED BRIDGES -

Land Use Variation on Mid-Columbia Plateau Upland and Lowland Archaeology Sites
Central Washington University ScholarWorks@CWU All Master's Theses Master's Theses Spring 2016 Land Use Variation on Mid-Columbia Plateau Upland and Lowland Archaeology Sites Cathy J. Anderson Central Washington University, [email protected] Follow this and additional works at: https://digitalcommons.cwu.edu/etd Part of the Anthropology Commons, and the Geography Commons Recommended Citation Anderson, Cathy J., "Land Use Variation on Mid-Columbia Plateau Upland and Lowland Archaeology Sites" (2016). All Master's Theses. 385. https://digitalcommons.cwu.edu/etd/385 This Thesis is brought to you for free and open access by the Master's Theses at ScholarWorks@CWU. It has been accepted for inclusion in All Master's Theses by an authorized administrator of ScholarWorks@CWU. For more information, please contact [email protected]. LAND USE VARIATION ON MID-COLUMBIA PLATEAU UPLAND AND LOWLAND ARCHAEOLOGY SITES A Thesis Presented to The Graduate Faculty Central Washington University In Partial Fulfillment of the Requirements for the Degree Master of Science Resource Management By Cathy Jean Anderson June 2016 i CENTRAL WASHINGTON UNIVERSITY Graduate Studies We here by approve the thesis of Cathy Jean Anderson Candidate for the degree of Master of Science APPROVED FOR THE GRADUATE FACULTY ______________ __________________________________________ Dr. Patrick T. McCutcheon, Committee Chair ______________ __________________________________________ Dr. Steven Hackenberger ______________ __________________________________________ Dr. Craig S. Revels ______________ __________________________________________ Dr. Kevin Archer, Dean of Graduate Studies ii ABSTRACT LAND USE VARIATION ON MID-COLUMBIA PLATEAU UPLAND AND LOWLAND ARCHAEOLOGY SITES by Cathy Jean Anderson June 9, 2016 Investigators of the Mid-Plateau archaeological record have interpreted artifact deposits in their environmental settings as evidence of human land use labeled as site types. -

Mountain Quail Translocations in Eastern Oregon
MOUNTAIN QUAIL TRANSLOCATIONS IN EASTERN OREGON Project Report: 2010 Trout Creek Mountains Kevyn Groot, Mountain Quail Technician Oregon Department of Fish and Wildlife, Hines District Office 237 Highway 20 South, P.O. Box 8 Hines, OR 97738 (541) 573-6582 1 Table of Contents Page Introduction……………………………………………………………………………………...2 Methods………………………………………………………………………………………….3 Results…………………………………………………………………………………………...6 Discussion………………………………………………………………………………….......12 Conclusion…………………………………………………………………………………..….17 Acknowledgements………………………………………………………………………....…18 References…………………………………………………………………………………..…19 Appendix I: Mountain Quail Movements from Release Site to Breeding Range by Project Year…………………………………………………………………….….21 Appendix II: List of Tables and Maps………………………………………………….…....22 Maps……………………………………………………………………………………………23 2 Introduction The primary distribution of the mountain quail (Oreortyx pictus) includes the Sierra Nevada, the Cascades, and the Coast Range of the western United States (Gutiérrez and Delehanty 1999). Populations in the western Great Basin are currently the focus of conservation efforts due to a steady decline that began in the 20th century (Crawford 2000). In particular, eastern Oregon, western Nevada, and western Idaho have shown evidence of diminished populations that warrant significant concern. Fire suppression, water development, and overgrazing by livestock in brush and riparian zones have contributed to the degradation of mountain quail habitat in these regions and are cited as the main causes of population declines (Brennan 1994 in Gutiérrez and Delehanty 1999, Pope and Crawford 2000). The Mountain Quail Translocation Program for Eastern Oregon began in 2001 as a cooperative effort between the Oregon Department of Fish and Wildlife (ODFW), the U.S. Forest Service, and the Game Bird Research Program at Oregon State University. The goal of this initiative is to actively restore viable, self-sustaining mountain quail populations to affected native ranges in eastern Oregon. -

High Desert Region Around Bend, Oregon by Lee Foster
High Desert Region Around Bend, Oregon by Lee Foster Beauty of nature in an alpine setting and diverse outdoor sports attract visitors to the Bend region of Central Oregon. Perusing natural beauty is the most universal pleasure here. Snow-capped mountains, pristine lakes, white-water rivers, and pine forests abound. At any time, the wilderness scenery is striking, with one of the dominant peaks, Mt. Bachelor, Broken Top, and the Sisters, usually present on your horizon. The main natural imprint on the land is a black volcanic presence. For the geology enthusiast, the Lava Lands Visitor Center explains the historic volcanic flows that form a stark legacy. Lava Butte is a 500-foot-high cinder cone, a silent reminder of past volcanic upheavals. A Rockhound Pow-Wow gathers amateur geologists here each July. Since opening in 1982, the High Desert Museum, south of Bend, has emerged as the most important nature interpretive effort in the state. (The High Desert Museum at Bend parallels Tucson’s Arizona-Sonora Desert Museum.) The raptor exhibit alone is worth the visit, putting you as close as you may ever get to a great horned owl, a red-tailed hawk, and an American kestrel. Foremost among the outdoor sports here is skiing at Mt. Bachelor. An extremely long ski season, both for alpine and nordic skiing, lasts into summer. The high- elevation chair lift to the top of Bachelor is popular also with non-skiers who seek an inspiring view of the region. In summer, hikers and campers depart from Bend for the nearby wilderness areas. -

Influence of Water Temperature and Beaver Ponds on Lahontan Cutthroat Trout in a High-Desert Stream, Southeastern Oregon
AN ABSTRACT OF THE THESIS OF Andrew G. Talabere for the degree of Master of Science in Fisheries Science presented on November 21. 2002. Title: Influence of Water Temperature and Beaver Ponds on Lahontan Cutthroat Trout in a High-Desert Stream, Southeastern Oregon Abstract approved Redacted for Privacy Redacted for Privacy The distribution of Lahontan cutthroat trout Oncorhynchus clarki henshawi was assessed in a high-desert stream in southeastern Oregon where beaver Castor canadensis are abundant. Longitudinal patterns of beaver ponds, habitat, temperature, and Lahontan cutthroat trout age group distribution were identified throughout Willow Creek. Three distinct stream segments were classified based on geomorphological characteristics. Four beaver-pond and four free-flowing sample sections were randomly located in each of the three stream segments. Beavers substantially altered the physical habitat of Willow Creek increasing the depth and width of available habitat. In contrast, there was no measurable effect on water temperature. The total number of Lahontan cutthroat trout per meter was significantly higher in beaver ponds than free-flowing sections. Although density (fish! m2) showed no statistically significant (P < 0.05) increase, values in beaver pondswere two-fold those of free-flowing sections. Age- 1 and young-of-the-year trout were absent or in very low numbers in lower Willow Creek because of elevated temperatures, but high numbers of age-2 and 3 (adults) Lahontan cutthroat trout were found in beaver ponds where water temperatures reached lethal levels (>24°C). Apparently survival is greater in beaver ponds than free-flowing sections as temperatures approach lethal limits. Influence of Water Temperature and Beaver Ponds on Lahontan Cutthroat Trout in a High- Desert Stream, Southeastern Oregon by Andrew G.