January-April 2021 Newsletter
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Message from the New Chairman
Subcommission on Devonian Stratigraphy Newsletter No. 21 April, 2005 MESSAGE FROM THE NEW CHAIRMAN Dear SDS Members: This new Newsletter gives me the pleasant opportunity to thank you for your confidence which should allow me to lead our Devonian Subcommission successfully through the next four years until the next International Geological Congress in Norway. Ahmed El Hassani, as Vice-Chairman, and John Marshall, as our new Secretary, will assist and help me. As it has been our habit in the past, our outgoing chairman, Pierre Bultynck, has continued his duties until the end of the calendar year, and in the name of all the Subcommission, I like to express our warmest thanks to him for all his efforts, his enthusi- asm for our tasks, his patience with the often too slow progress of research, and for the humorous, well organized and skil- ful handling of our affairs, including our annual meetings. At the same time I like to thank all our outgoing Titular Members for their partly long-time service and I express my hope that they will continue their SDS work with the same interest and energy as Corresponding Members. The new ICS rules require a rather constant change of voting members and the change from TM to CM status should not necessarily be taken as an excuse to adopt the lifestyle of a “Devonian pensioner”. I see no reason why constantly active SDS members shouldn´t become TM again, at a later stage. On the other side, the rather strong exchange of voting members should bring in some fresh ideas and some shift towards modern stratigraphical tech- niques. -

001-012 Primeras Páginas
PUBLICACIONES DEL INSTITUTO GEOLÓGICO Y MINERO DE ESPAÑA Serie: CUADERNOS DEL MUSEO GEOMINERO. Nº 9 ADVANCES IN TRILOBITE RESEARCH ADVANCES IN TRILOBITE RESEARCH IN ADVANCES ADVANCES IN TRILOBITE RESEARCH IN ADVANCES planeta tierra Editors: I. Rábano, R. Gozalo and Ciencias de la Tierra para la Sociedad D. García-Bellido 9 788478 407590 MINISTERIO MINISTERIO DE CIENCIA DE CIENCIA E INNOVACIÓN E INNOVACIÓN ADVANCES IN TRILOBITE RESEARCH Editors: I. Rábano, R. Gozalo and D. García-Bellido Instituto Geológico y Minero de España Madrid, 2008 Serie: CUADERNOS DEL MUSEO GEOMINERO, Nº 9 INTERNATIONAL TRILOBITE CONFERENCE (4. 2008. Toledo) Advances in trilobite research: Fourth International Trilobite Conference, Toledo, June,16-24, 2008 / I. Rábano, R. Gozalo and D. García-Bellido, eds.- Madrid: Instituto Geológico y Minero de España, 2008. 448 pgs; ils; 24 cm .- (Cuadernos del Museo Geominero; 9) ISBN 978-84-7840-759-0 1. Fauna trilobites. 2. Congreso. I. Instituto Geológico y Minero de España, ed. II. Rábano,I., ed. III Gozalo, R., ed. IV. García-Bellido, D., ed. 562 All rights reserved. No part of this publication may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopy, recording, or any information storage and retrieval system now known or to be invented, without permission in writing from the publisher. References to this volume: It is suggested that either of the following alternatives should be used for future bibliographic references to the whole or part of this volume: Rábano, I., Gozalo, R. and García-Bellido, D. (eds.) 2008. Advances in trilobite research. Cuadernos del Museo Geominero, 9. -

Eurypterids Xenacanthus
animals of pennsylvania Did you know that there are many types of animals (other than dinosaurs) that paleontologists study? Pennsylvania had many diverse and interesting pre-historic animals. Some lived thousands of years ago, but some lived millions of years ago. Check out these pre-historic animals from Pennsylvania’s past! 1 Eurypterids A relative to our modern-day horseshoe crab, sea scorpions thrived in the oceans, brackish, and freshwater during the Ordovician through Permian Periods with their peak being during the Silurian Period (443.8 million years ago to 419.2 million years ago). They were arthropods, invertebrate animals with a hard exoskeleton, segmented body, and paired jointed appendages. Today’s arthropods include not only horseshoe crabs, but Image Credit: Jaime Chirinos also insects, spiders, crabs, and today’s land-dwelling scorpions. Most eurypterids were 8 inches in length or smaller, but varieties up to 8.2 feet long did exist. Most of Pennsylvania’s eurypterid fossils are around 1 foot long or less in size. They were some of the largest predators living during their time. One of the modern shark’s early ancestors is xenacanthus which lived in Pennsylvania from 2 Xenacanthus the Late Devonian through Triassic Period (400 to 202 million years ago). Xenacanthus lived in freshwater and ranged from 3-6 feet in length. With the possibly venomous head spike giving it its name, the Unicorn Shark’s back fin was long and ran down the entire length of the spine. Being cartilaginous (mostly made of soft tissue, like that in your nose and ears), like the modern shark, xenacanthus has been identified from its fossilized teeth and spines. -

Torney, Clare (2011) Mineral Eyes : Lessons from the Natural World. Phd Thesis
Torney, Clare (2011) Mineral eyes : lessons from the natural world. PhD thesis. http://theses.gla.ac.uk/2331/ Copyright and moral rights for this thesis are retained by the author A copy can be downloaded for personal non-commercial research or study, without prior permission or charge This thesis cannot be reproduced or quoted extensively from without first obtaining permission in writing from the Author The content must not be changed in any way or sold commercially in any format or medium without the formal permission of the Author When referring to this work, full bibliographic details including the author, title, awarding institution and date of the thesis must be given Glasgow Theses Service http://theses.gla.ac.uk/ [email protected] Mineral Eyes – Lessons from the Natural World Clare Torney BSc. (Hons) University of Glasgow Submitted in fulfilment of the requirements for the Degree of Doctor of Philosophy School of Geographical and Earth Sciences College of Science and Engineering University of Glasgow September 2010 ii Abstract The compound eyes of trilobites, which appeared in the Early Cambrian, represent one of the first preserved visual systems. Application of state-of-the- art microscopy techniques in the present study has revealed fine details of the microstructure and chemistry of these unusual calcite eyes that, until now, have been inaccessible and this has facilitated new insights into their growth and function. Six species from three families of trilobite with holochroal eyes, ranging from Early Ordovician to Middle Carboniferous, and 21 species from three families of trilobite with schizochroal eyes, ranging from Early Ordovician to Middle Devonian, were investigated. -

Fossil Catalog #29
GEOLOGICAL ENTERPRISES P.O. BOX 996 -- ARDMORE, OKLAHOMA 73402 USA phone 580-223-8537 fax 580-223-6965 email [email protected] website WWW.GEOLOGICALENTERPRISES.COM FOSSIL CATALOG #29 GEOLOGICAL ENTERPRISES, INC. P.O. BOX 996 ARDMORE, OKLAHOMA U.S.A. 73402 Phone 580-223-8537 Fax 580-223-6965 email [email protected] VISIT US ON THE WEB @ www.geologicalenterprises.com PLEASE TAKE CARE OF THIS CATALOG: Due to the high costs of printing and mailing, we will only issue a new catalog periodically. We literally send out thousands of these to Universities worldwide at no charge. These catalogs are used by Professors and students, for reference, and in some cases, as text books. We will continue to issue our yearly bulletin, which will contain additions to this catalog. We pledge to keep the quality of the specimens listed here as high as possible. We attempt to secure the finest available specimens at all times. Complete geological data accompanies all specimens. TERMS: We accept VISA, MasterCard, Discover Card, Personal or Company Checks, Money Orders and Pay Pal. (Please use our regular email address [email protected] for PayPal) To recognized educational and corporate institutions, our terms are Net-30 days. All other orders should be prepaid. We accept only checks drawn on U.S. Banks. Wire transfers are also acceptable. Please be aware Wire transfers may incur a fee. All prices are F.O.B. Ardmore, Oklahoma. Please allow for postage(5% is usually sufficient. $8.00 minimum). Overpayments will be promptly refunded. WEBSITE: Our Catalog is also available on our website in color! It’s truly beautiful. -
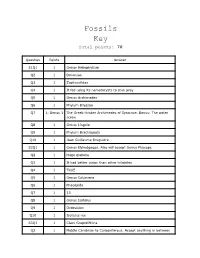
Fossils Key Total Points: 70
Fossils Key Total points: 70 Question Points Answer S1Q1 1 Genus Heliophyllum Q2 1 Devonian Q3 1 Zaphrentidae Q4 1 It fed using its nematocysts to stun prey Q5 1 Genus Archimedes Q6 1 Phylum Bryozoa Q7 1; Bonus 1 The Greek thinker Archimedes of Syracuse. Bonus: The water screw Q8 1 Genus Lingula Q9 1 Phylum Brachiopoda Q10 1 Jean Guillaume Bruguière S2Q1 1 Genus Eldredgeops. Also will accept Genus Phacops Q2 1 Huge glabella Q3 1 It had better vision than other trilobites Q4 1 TRUE Q5 1 Genus Calymene Q6 1 Phacopida Q7 1 13 Q8 1 Genus Isotelus Q9 1 Ordovician Q10 1 Isotelus rex S3Q1 1 Class Graptolithina Q2 1 Middle Cambrian to Carboniferous. Accept anything in between Q3 1 Accept graptos or lithos Q4 1 Phylum Hemichordata Q5 1 Genus Gryphaea Q6 1 Devil’s toenails Q7 1 TRUE Q8 1 Genus Dactylioceras Q9 1 Subclass Ammonoidea (Ammonoids) Q10 1 dactyl, meaning finger S4Q1 1 Genus Bothriolepis Q2 1 Bothriolepididae Q3 1 That external nasal openings lay on the dorsal side of the head near the eyes Q4 1 They break down easily and decompose much faster than hard tissues Q5 1 Genus Acanthostega Q6 1 Recognizable limbs Q7 1 TRUE Q8 1 Order Coelacanthiformes Q9 1 Class Sarcopterygii Q10 1 TRUE S5Q1 1 Genus Diplodocus Q2 1 1877 Q3 1 Othniel Charles Marsh Q4 1 Its forelimbs are slightly shorter than its hind limbs Q5 1 Genus Tyrannosaurus Q6 1 Order Saurischia Q7 1 Western North America. Will also accept North America Q8 1 Genus Dracorex Q9 1 Dracorex hogwartsia Q10 1 dragon king of Hogwarts S6Q1 1 Species neanderthalensis Q2 1 Eurasia Q3 1 Any of the following: neanderthals are stockier, had shorter legs, had bigger bodies Q4 1 Genus Dimetrodon Q5 1 Within 50 Ma: 295-272 Ma Q6 1 Sphenacodontidae Q7 1 Genus Smilodon Q8 1 Class Mammalia Q9 1 Pleistocene epoch Q10 1 1842 S7Q1 1 Shale Q2 1 Compaction Q3 1 Clastic Q4 1 Sandstone Q5 1 Any of the following: river, ocean, glacier, desert deposits Q6 1 clastic Q7 1 Coquina Q8 1 FALSE Q9 1 Any of the following: shells of molluscs, trilobites, brachiopods, or other invertebrates Q10 1 organic. -

A Window Into the Devonian Period of Western New York
PENN DIXIE FOSSIL PARK & NATURE RESERVE: A WINDOW INTO THE DEVONIAN PERIOD OF WESTERN NEW YORK Philip J. Stokes, PhD and Holly A. Schreiber, PhD Penn Dixie Fossil Park & Nature Reserve, 3556 Lakeshore Road, Suite 230, Blasdell, NY 14219 E-mail: [email protected]; [email protected] An Image Caught in Time – Earth was very different four hundred million years ago (Figure 1). Eons before the first dinosaurs scuttled through prehistoric forests, North America and Europe together formed a large landmass along the Tropic of Capricorn (Figure 2B). Western New York was submerged beneath tropical seas, which were teeming with both old and new forms of life. The Devonian Period (419 to 359 million years ago) marked an important chapter in Earth history. Life on land was just starting to take root — literally, as Earth’s first forests produced the oxygen needed by the earliest amphibians who hunted giant insects at the water’s edge. Along the Eastern half of North America, mountains rose as the Acadian Orogeny (i.e., mountain building event) -- was underway. Like a highway pileup, the Proto-African Plate crashed into and onto the North American plate, causing earthquakes, intense volcanic activity, and crustal deformation (Figure 2C). Beginning in the middle of the Devonian Period and lasting for 60 million years, The Acadian Orogeny set the stage for our unique assemblage of fossils. To the West of the coastal collision zone, the Earth’s crust flexed and bent to accommodate the incoming mass of another continent. Like a compressed accordion, land was forced downwards. Over centuries, the ocean meandered its way along the widening and deepening depression towards New York. -

University of Cincinnati
! "# $ % & % ' % ! !' " # $$ %&'& ( "#) *# +, ( !, &$ ' "# ' '% $$(' - , -. ' #"! " $ /0& $ Anatomy of Middle Devonian Faunal Turnover in Eastern North America: Implications for Global Bioevents at the Eifelian-Givetian Stage Boundary A dissertation submitted to the Graduate School University of Cincinnati in partial fulfilment of the requirements for the degree of Doctor of Philosophy Department of Geology McMicken College of Arts and Sciences November 2010 By Michael K. DeSantis B.A., Boston University, 1992 M.S., University of Idaho, 1996 Committee Chair: Carlton E. Brett, Ph.D. ABSTRACT The late Eifelian-earliest Givetian interval (Middle Devonian) represents a time of significant faunal turnover both in the eastern North America and globally, herein termed the Late Eifelian Biocrisis. A synthesis of biostratigraphic, K-bentonite, and sequence stratigraphic data, indicates that physical and biotic events in the Appalachian foreland basin sections in New York are coeval with the predominantly carbonate platform sections of the adjacent midcontinent. The succession includes portions of three distinctive benthic faunas or ecological-evolutionary sub- units (EESUs): ‘Onondaga’, ‘lower Marcellus’, and ‘Hamilton’. In the northern Appalachian Basin in New York the boundaries of these bioevents show evidence of abrupt, widespread extinctions/extirpations, immigration and ecological restructuring. Regional analysis of faunas shows that EESUs comparable to the northern Appalachian Basin succession are recognizable within the coeval carbonate ramp and platform strata of central Ohio and southeastern Indiana, as well as in the Michigan Basin. However, faunas of the Wabash Platform area of central Ohio and southeastern Indiana show slightly higher persistence compared to those of the Appalachian Basin, indicating that the Wabash Platform served as a refuge for species extirpated from the Appalachian Basin by environmental changes brought on by tectonic and eustatic deepening. -

Anatomy & Physiology
ANATOMY & PHYSIOLOGY See General Rules, Eye Protection & other Policies on www.soinc.org as they apply to every event. 1. DESCRIPTION: Participants will be assessed on their understanding of the anatomy and physiology for the human Cardiovascular, Lymphatic, and Excretory systems. A TEAM OF UP TO: 2 APPROXIMATE TIME: 50 Minutes 2. EVENT PARAMETERS: Each team may bring one 8.5” x 11” sheet of paper that may contain information on both sides in any form and from any source along with two stand-alone non-programmable, non-graphing calculators. 3. THE COMPETITION: Participants will complete a written test limited to the following topics. Topics listed in italics will only be assessed at the National Tournament. a. CARDIOVASCULAR SYSTEM: i. Anatomy and physiology of the cardiovascular system ii. The Heart - chambers and valves of the heart, electrical stimulation of myocardial tissue, pacemaker tissue, interpreting ECG (EKG) readings on strips iii. Blood Vessels – structure and function of arteries, arterioles, veins, venules, capillaries, including the functionality of Startling’s forces in the capillaries iv. Blood - plasma, hematocrit, red blood cells, oxygen transport, hemoglobin and cooperative binding of oxygen, platelets and blood clotting, regulation of blood plasma volume and acidity, blood typing & basic genetics of ABO, Rh, blood types v. Measurement of the pulse rate and blood pressure with appropriate instrumentation vi. Calculations include systolic and diastolic pressure, mean arterial pressure, stroke volume & cardiac output vii. Effects of exercise, smoking, alcohol, caffeine, and drugs on the cardiovascular system viii. Understand disorders: Congestive Heart Failure, Atrial Fibrillation, Myocardial Infarction, Atherosclerosis, Bradycardia, and Tachycardia ix. -

The Late Middle Devonian (Givetian) Global Taghanic Biocrisis in Its
The Late Middle Devonian (Givetian) Global Taghanic Biocrisis in its Type Region (Northern Appalachian Basin): Geologically Rapid Faunal Transitions Driven by Global and Local Environmental Changes A dissertation submitted to the Graduate School University of Cincinnati In partial fulfillment of the requirements for the degree of DOCTOR OF PHILOSOPHY Department of Geology McMicken College of Arts and Sciences August 9th, 2011 by James Joseph Zambito IV M.S., University at Buffalo, 2006 B.S., SUNY College at Brockport, 2004 Dissertation Committee Dr. Carlton E. Brett, Chair Dr. Arnold I. Miller Dr. David L. Meyer Dr. Thomas J. Algeo Dr. Gordon C. Baird Dr. Alex J. Bartholomew Abstract The late Middle Devonian Global Taghanic Biocrisis marks the onset of extinction and a loss of faunal endemism that culminated in the subsequent Frasnian-Famennian extinction. Global environmental changes recognized at this time include increased warming and aridity, as well as rapid eustatic sea level fluctuations. In the type region, the northern Appalachian Basin, the biocrisis is recorded within the deposits of the uppermost Hamilton, Tully, and lowermost Genesee Groups over an interval of ~0.5 million years. A high-resolution stratigraphic framework reconstructed along a complete onshore through offshore gradient has resulted in the recognition of three main pulses (bioevents) of faunal transition in the type region: 1) the incursion of the tropical Tully Fauna into eastern Laurentia and temporary loss of the endemic Hamilton Fauna; 2) Tully Fauna extermination and replacement by a recurrent Hamilton Fauna; and, ultimately, 3) extinction of large portions of the Hamilton Fauna and the beginning of cosmopolitan Genesee Fauna. -

The Eyes of Trilobites: the Oldest Preserved Visual System
Arthropod Structure & Development 35 (2006) 247e259 www.elsevier.com/locate/asd Review The eyes of trilobites: The oldest preserved visual system Euan Clarkson a,*, Riccardo Levi-Setti b, Gabor Horva´th c a Grant Institute of Earth Sciences, School of Geosciences, University of Edinburgh, Kings Buildings, West Mains Road, Edinburgh EH9 3JW, UK b Enrico Fermi Institute and Department of Physics, University of Chicago, 5630 Ellis Avenue, Chicago, IL 10024, USA c Biooptics Laboratory, Department of Biological Physics, Lorand Eo¨tvo¨s University, Pazmany Peter setany 1, H-1117 Budapest, Hungary Received 22 April 2006; accepted 1 August 2006 Abstract The oldest preserved visual systems are to be found in the extinct trilobites, marine euarthropods which existed between about 520 and 250 million years ago. Because they possessed a calcified cuticle, they have a good fossil record, and commonly the lens-bearing surfaces of their paired compound eyes are well preserved. The sublensar structures, however, remain unknown. Three kinds of eyes have been distin- guished. Holochroal eyes, apomorphic for trilobites, typically have many contiguous small lenses, set on a kidney-shaped visual surface. Lens optics, angular range of vision, and ontogeny have been established for many compound eyes. Some pelagic trilobites have enormous eyes, subtending a panoramic field of view. Schizochroal eyes are found only in one group, the phacopids (Ordovician to Devonian). These have large lenses, separated from each other by cuticular material, and the lenses have a complex doublet or triplet internal structure, which could focus light sharply. The optics of phacopid eyes are becoming increasingly well known despite the fact that there are no direct counterparts in any living arthropods today. -

Abstract Volume, and the Entire Conference, Is Dedicated to the Memory of Professor Harry Blackmore Whittington FRS (March 24, 1916 – June 20, 2010)
CZECH GEOLOGICAL SURVEY & CHARLES UNIVERSITY, PRAGUE CZECH GEOLOGICAL SURVEY & CHARLES UNIVERSITY, PRAGUE The th5 Conference on Trilobites and their relatives 1st July – 4th July 2012, Prague, Czech Republic ABSTRACTS Editors: Petr Budil & Oldřich Fatka ISBN: 978-80-7075-787-1 The th5 Conference on Trilobites and their relatives ABSTRACTS Cover illustration: Conocoryphe sulzeri sulzeri (Schlotheim, 1823). Entire specimen (CGS VV 3) with healed injury on the right gena. “Middle” Cambrian, Drumian, Jince Formation, Paradoxides gracilis Biozone. Locality Rejkovice-Řešátko. Leg. L. Grigar, 1999. The th5 Conference on Trilobites and their relatives 1st July – 4th July 2012, Prague, Czech Republic ABSTRACTS Editors: Petr Budil & Oldřich Fatka Czech Geological Survey & Charles University, Prague Organising committee: Oldřich Fatka (Charles University, Prague) Petr Budil (Czech Geological Survey, Prague) Gian luiGi PilOlla (University of Cagliari, Italy) Michal MerGl (West Bohemian University, Plzeň) ŠtěPán rak (Museum of Bohemian Karst, Beroun) Martin Valent (National Museum, Prague) radkO Šarič (Czech Geological Survey, Prague) Marika SteinOVá (Czech Geological Survey, Prague) Scientific committee: P. ahlBerG (Lund University, Sweden) B.d.e. chattertOn (University of Alberta, Canada) c. crOnier (University Lille, France) r.a. FOrtey (Natural History Museum, London, Britain) d. hOllOway (Museum Victoria, Melbourne, Australia) r. lerOSey-auBril (Forschungsinstitut Senckenberg, Frankfurt a/M, Germany) e. naiMark (Russian Academy of Sciences, Moscow,