2016 일반생물학 Ch.16 Prokaryote
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Morphology, Phylogeny, and Diversity of Trichonympha (Parabasalia: Hypermastigida) of the Wood-Feeding Cockroach Cryptocercus Punctulatus
J. Eukaryot. Microbiol., 56(4), 2009 pp. 305–313 r 2009 The Author(s) Journal compilation r 2009 by the International Society of Protistologists DOI: 10.1111/j.1550-7408.2009.00406.x Morphology, Phylogeny, and Diversity of Trichonympha (Parabasalia: Hypermastigida) of the Wood-Feeding Cockroach Cryptocercus punctulatus KEVIN J. CARPENTER, LAWRENCE CHOW and PATRICK J. KEELING Canadian Institute for Advanced Research, Botany Department, University of British Columbia, University Boulevard, Vancouver, BC, Canada V6T 1Z4 ABSTRACT. Trichonympha is one of the most complex and visually striking of the hypermastigote parabasalids—a group of anaerobic flagellates found exclusively in hindguts of lower termites and the wood-feeding cockroach Cryptocercus—but it is one of only two genera common to both groups of insects. We investigated Trichonympha of Cryptocercus using light and electron microscopy (scanning and transmission), as well as molecular phylogeny, to gain a better understanding of its morphology, diversity, and evolution. Microscopy reveals numerous new features, such as previously undetected bacterial surface symbionts, adhesion of post-rostral flagella, and a dis- tinctive frilled operculum. We also sequenced small subunit rRNA gene from manually isolated species, and carried out an environmental polymerase chain reaction (PCR) survey of Trichonympha diversity, all of which strongly supports monophyly of Trichonympha from Cryptocercus to the exclusion of those sampled from termites. Bayesian and distance methods support a relationship between Tricho- nympha species from termites and Cryptocercus, although likelihood analysis allies the latter with Eucomonymphidae. A monophyletic Trichonympha is of great interest because recent evidence supports a sister relationship between Cryptocercus and termites, suggesting Trichonympha predates the Cryptocercus-termite divergence. -

Biology of Protists Video and DVD Guide. the BIOLOGY of PROTISTS Produced by Biomedia ASSOCIATES ©2003 -- Running Time 20 Minutes
Biology of Protists video and DVD guide. THE BIOLOGY OF PROTISTS Produced by BioMEDIA ASSOCIATES ©2003 -- Running time 20 minutes. Order Toll Free (877) 661-5355 or FAX (843) 470-0237 Or mail orders to eBioMEDIA, P.O. Box 1234, Beaufort, SC 29901–1234 (IMAGES IN THIS GUIDE ARE FROM THE VIDEO PROGRAM) Cosmerium, a green alga Arcella, a shelled ameoba The goal of this program is to show a representative sample of the great diversity of protists, and to show why they need a new classification reflecting our growing understanding of their long evolutionary history. The protists shown can be found in habitats such as: roadside puddles, park duck ponds, aquariums, birdbaths and in the gut of termites. We hope that these observations will encourage students to collect pond water samples and see for themselves this amazing hidden world. The live photography was accomplished using a variety microscope techniques, including dark field (good for showing the natural color of subjects) and the most advanced forms of differential interference contrast (DIC gives contrast to transparent structures that would be invisible in normal bright field microscopy). While these forms of imaging living protists are particularly revealing for some aspects of micro-organism biology, all of the organisms seen here can be studied successfully with student microscopes. IMPORTANT NOTE: This program packs a lot of information and a wealth of observational data into nine different modules. We recommend that the material be viewed module by module, stopping for discussion and replay as needed. “Discussion starters” for the various modules are provided in this guide. -

A Resurgence in Field Research Is Essential to Better Understand
Received Date : 15-Mar-2013 Revised Date : 21-Oct-2013 Accepted Date : 29-Oct-2013 Article type : Symposium Article Heger et al. --- Importance of Field Protistology A Resurgence in Field Research is Essential to Better Understand the Diversity, Ecology, and Evolution of Microbial Eukaryotes Thierry J. Hegera, Virginia P. Edgcombb, Eunsoo Kimc, Julius Lukešd, Brian S. Leandera & Naoji Yubukia a The Departments of Botany and Zoology, Beaty Biodiversity Research Centre and Museum, University of British Columbia, Vancouver, British Columbia, V6T 1Z4, Canada b Woods Hole Oceanographic Institution, Geology and Geophysics Department, Woods Hole, Article MA 02543, USA c Division of Invertebrate Zoology, American Museum of Natural History, New York, NY, 10024, USA d Institute of Parasitology, Biology Centre, Czech Academy of Sciences, and Faculty of Science, University of South Bohemia, 37005 České Budějovice, Czech Republic Correspondence T. J. Heger and N. Yubuki, The Departments of Botany and Zoology, Beaty Biodiversity Research Centre and Museum, University of British Columbia, 6270 University Blvd., Vancouver, British Columbia, V6T 1Z4, Canada Telephone number: +1 604 822 4892; FAX number: +1 604 822 6089; emails: [email protected] and [email protected] ABSTRACT The discovery and characterization of protist communities from diverse environments are crucial for understanding the overall evolutionary history of life on earth. However, major questions about the diversity, ecology, and evolutionary history of protists remain unanswered, -
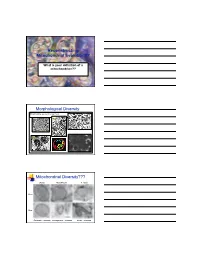
Reconstructing Mitochondrial Evolution?? Morphological Diversity Mitochondrial Diversity???
Reconstructing Mitochondrial Evolution?? What is your definition of a mitochondrion?? Morphological Diversity Mitochondria as we all know them: Suprarenal gland Liver cell Plasma cell Adrenal cortex Mitochondrial Diversity??? Chicken Neocallimastix T. foetus 100 nm 50 nm Entamoeba - mitosome microsporidian - mitosome Giardia - mitosome Reconstructing Evolution Mitochondrial evolution well established endosymbiotic theory α-proteobacterium - Rickettsia prowazekii Hydrogenosomal evolution No DNA NOW 2 examples Nyctotherus and Blastocystis (MLO) Several proteins similar to mitochondria Mitosome evolution No DNA Few proteins identified similar to mitochondria Origins via Endosymbiosis Current dogma - mitochondria and related Aerobic α-proteobacterium prokaryote gave rise to present day mitochondria. organelles arose just once in evolution Are hydrogenosomes and mitosomes of anaerobic protists derived from the same proto-mitochondrion? Evidence for: accumulating evidence for several proteins that are currently found in mitochondria - Proteins of Fe-S cluster formation. Scenario A Common ancestor facultative Scenario B organism? degenerate mitochondrion invoke lateral gene transfer from anaerobic prokaryotes Hypotheses for Mito Acquisition Mitochondrion-related Organelles 2 way to interpret this summary - can you think of both? Groups with mitochondrial homologues Hjort et al Phil. Trans. R. Soc. B 2010 365, 713-727 Origin of Hydrogenosomes (Mitosomes?) 1. a)Conversion of Mitochondria HYD b)Common ancestor with mitochondria ? HYD 2. Independent Origin α-proteobacterium ? - anaerobic bacterium HYD Organelles - origins and biogenesis Approaches: (1) Conduct phylogenetic analyses of similar proteins Hsp70 Fd Hsp60 Isc subunits (2) Examine protein targeting to the organelle matrix protein targeting membrane protein targeting (3) Characterize membrane/translocation components These components could have evolved as the endosymbiont was converted to organelle. Reveals evolutionary history. -

Superorganisms of the Protist Kingdom: a New Level of Biological Organization
Foundations of Science https://doi.org/10.1007/s10699-020-09688-8 Superorganisms of the Protist Kingdom: A New Level of Biological Organization Łukasz Lamża1 © The Author(s) 2020 Abstract The concept of superorganism has a mixed reputation in biology—for some it is a conveni- ent way of discussing supra-organismal levels of organization, and for others, little more than a poetic metaphor. Here, I show that a considerable step forward in the understand- ing of superorganisms results from a thorough review of the supra-organismal levels of organization now known to exist among the “unicellular” protists. Limiting the discussion to protists has enormous advantages: their bodies are very well studied and relatively sim- ple (as compared to humans or termites, two standard examples in most discussions about superorganisms), and they exhibit an enormous diversity of anatomies and lifestyles. This allows for unprecedented resolution in describing forms of supra-organismal organiza- tion. Here, four criteria are used to diferentiate loose, incidental associations of hosts with their microbiota from “actual” superorganisms: (1) obligatory character, (2) specifc spatial localization of microbiota, (3) presence of attachment structures and (4) signs of co-evolu- tion in phylogenetic analyses. Three groups—that have never before been described in the philosophical literature—merit special attention: Symbiontida (also called Postgaardea), Oxymonadida and Parabasalia. Specifcally, it is argued that in certain cases—for Bihos- pites bacati and Calkinsia aureus (symbiontids), Streblomastix strix (an oxymonad), Joe- nia annectens and Mixotricha paradoxa (parabasalids) and Kentrophoros (a ciliate)—it is fully appropriate to describe the whole protist-microbiota assocation as a single organism (“superorganism”) and its elements as “tissues” or, arguably, even “organs”. -

Bacterial and Archaeal Symbioses with Protists, Current Biology (2021), J.Cub.2021.05.049
Please cite this article in press as: Husnik et al., Bacterial and archaeal symbioses with protists, Current Biology (2021), https://doi.org/10.1016/ j.cub.2021.05.049 ll Review Bacterial and archaeal symbioses with protists Filip Husnik1,2,*, Daria Tashyreva3, Vittorio Boscaro2, Emma E. George2, Julius Lukes3,4, and Patrick J. Keeling2,* 1Okinawa Institute of Science and Technology, Okinawa, 904-0495, Japan 2Department of Botany, University of British Columbia, Vancouver, V6T 1Z4, Canada 3Institute of Parasitology, Biology Centre, Czech Academy of Sciences, Ceske Budejovice, 370 05, Czech Republic 4Faculty of Science, University of South Bohemia, Ceske Budejovice, 370 05, Czech Republic *Correspondence: fi[email protected] (F.H.), [email protected] (P.J.K.) https://doi.org/10.1016/j.cub.2021.05.049 SUMMARY Most of the genetic, cellular, and biochemical diversity of life rests within single-celled organisms—the pro- karyotes (bacteria and archaea) and microbial eukaryotes (protists). Very close interactions, or symbioses, between protists and prokaryotes are ubiquitous, ecologically significant, and date back at least two billion years ago to the origin of mitochondria. However, most of our knowledge about the evolution and functions of eukaryotic symbioses comes from the study of animal hosts, which represent only a small subset of eukary- otic diversity. Here, we take a broad view of bacterial and archaeal symbioses with protist hosts, focusing on their evolution, ecology, and cell biology, and also explore what functions (if any) the symbionts provide to their hosts. With the immense diversity of protist symbioses starting to come into focus, we can now begin to see how these systems will impact symbiosis theory more broadly. -

Trichonympha Cf
MOLECULAR PHYLOGENETICS OF TRICHONYMPHA CF. COLLARIS AND A PUTATIVE PYRSONYMPHID: THE RELEVANCE TO THE ORIGIN OF SEX by JOEL BRYAN DACKS B.Sc. The University of Alberta, 1995 A THESIS SUBMITTED IN PARTIAL FULFILMENT OF THE REQUIREMENTS FOR THE DEGREE OF MASTER'S OF SCIENCE in THE FACULTY OF GRADUATE STUDIES (Department of Zoology) We accept this thesis as conforming to the required standard THE UNIVERSITY OF BRITISH COLUMBIA April 1998 © Joel Bryan Dacks, 1998 In presenting this thesis in partial fulfilment of the requirements for an advanced degree at the University of British Columbia, I agree that the Library shall make it freely available for reference and study. I further agree that permission for extensive copying of this thesis for scholarly purposes may be granted by the head of my department or by his or her representatives. It is understood that copying or publication of this thesis for financial gain shall not be allowed without my written permission. Department of ~2—oc)^Oa^ The University of British Columbia Vancouver, Canada Date {X^ZY Z- V. /^P DE-6 (2/88) Abstract Why sex evolved is one of the central questions in evolutionary genetics. To address this question I have undertaken a molecular phylogenetic study of two candidate lineages to determine the first sexual line. In my thesis the hypermastigotes are confirmed as closely related to the trichomonads in the phylum Parabasalia and found to be more deeply divergent than a putative pyrsonymphid. This means that the Parabasalia are the first sexual lineage. From this I go on to infer that the ancestral sexual cycle included facultative sex. -

The Amoeboid Parabasalid Flagellate Gigantomonas Herculeaof
Acta Protozool. (2005) 44: 189 - 199 The Amoeboid Parabasalid Flagellate Gigantomonas herculea of the African Termite Hodotermes mossambicus Reinvestigated Using Immunological and Ultrastructural Techniques Guy BRUGEROLLE Biologie des Protistes, UMR 6023, CNRS and Université Blaise Pascal de Clermont-Ferrand, Aubière Cedex, France Summary. The amoeboid form of Gigantomonas herculea (Dogiel 1916, Kirby 1946), a symbiotic flagellate of the grass-eating subterranean termite Hodotermes mossambicus from East Africa, is observed by light, immunofluorescence and transmission electron microscopy. Amoeboid cells display a hyaline margin and a central granular area containing the nucleus, the internalized flagellar apparatus, and organelles such as Golgi bodies, hydrogenosomes, and food vacuoles with bacteria or wood particles. Immunofluorescence microscopy using monoclonal antibodies raised against Trichomonas vaginalis cytoskeleton, such as the anti-tubulin IG10, reveals the three long anteriorly-directed flagella, and the axostyle folded into the cytoplasm. A second antibody, 4E5, decorates the conspicuous crescent-shaped structure or cresta bordered by the adhering recurrent flagellum. Transmission electron micrographs show a microfibrillar network in the cytoplasmic margin and internal bundles of microfilaments similar to those of lobose amoebae that are indicative of cytoplasmic streaming. They also confirm the internalization of the flagella. The arrangement of basal bodies and fibre appendages, and the axostyle composed of a rolled sheet of microtubules are very close to that of the devescovinids Foaina and Devescovina. The very large microfibrillar cresta supporting an enlarged recurrent flagellum resembles that of Macrotrichomonas. The parabasal apparatus attached to the basal bodies is small in comparison to the cell size; this is probably related to the presence of many Golgi bodies supported by a striated fibre that are spread throughout the central cytoplasm in a similar way to Placojoenia and Mixotricha. -

Parasitology Mgr. Anna Novák Vanclová Evolution of Euglenid Plastid Proteome
Charles University, Faculty of Science Univerzita Karlova, Přírodovědecká fakulta Study program: Parasitology Studijní program: Parazitologie Mgr. Anna Novák Vanclová Evolution of euglenid plastid proteome Evoluce proteomu plastidu euglenidů Summary of the doctoral thesis Thesis supervisor: doc. Vladimír Hampl, Ph.D. Prague, 2019 ABSTRACT Endosymbiotic gain and transfer of plastids is a widespread evolutionary phenomenon and a major driving force of eukaryotic evolution. The integration of a new organelle is accompanied by changes in its structure, gene content, molecular mechanisms for biogenesis and transport, and re-wiring of the host and organelle metabolic pathways. To understand the course and underlying mechanisms of plastid evolution, it is important to study these processes in variety of secondary algae and notice their differences and similarities. Euglenophytes gained their plastids from green eukaryotic algae after a long history of heterotrophic lifestyle. In my thesis, I participated in analyses of newly generated sequence datasets: transcriptomes of Euglena gracilis and Euglena longa and mass spectrometry-determined proteome of E. gracilis plastid with especial regard to the potential novelties associated with plastid gain and incorporation. In the resulting publications we particularly focus on plastid protein import machinery and targeting signals and report extremely reduced TIC and completely absent TOC in euglenophyte plastid. Using the proteomic dataset, we predict potential novel plastid protein translocases recruited from ER/Golgi and re-analyze plastid signal domains, characterizing previously overlooked features. Protein inventory of E. gracilis plastid suggests complex, in some cases redundant metabolic capacity. Chlorophyll recycling is one of the sources of phytol for reactions not connected to MEP/DOXP pathway. -

Catalogue of Protozoan Parasites Recorded in Australia Peter J. O
1 CATALOGUE OF PROTOZOAN PARASITES RECORDED IN AUSTRALIA PETER J. O’DONOGHUE & ROBERT D. ADLARD O’Donoghue, P.J. & Adlard, R.D. 2000 02 29: Catalogue of protozoan parasites recorded in Australia. Memoirs of the Queensland Museum 45(1):1-164. Brisbane. ISSN 0079-8835. Published reports of protozoan species from Australian animals have been compiled into a host- parasite checklist, a parasite-host checklist and a cross-referenced bibliography. Protozoa listed include parasites, commensals and symbionts but free-living species have been excluded. Over 590 protozoan species are listed including amoebae, flagellates, ciliates and ‘sporozoa’ (the latter comprising apicomplexans, microsporans, myxozoans, haplosporidians and paramyxeans). Organisms are recorded in association with some 520 hosts including mammals, marsupials, birds, reptiles, amphibians, fish and invertebrates. Information has been abstracted from over 1,270 scientific publications predating 1999 and all records include taxonomic authorities, synonyms, common names, sites of infection within hosts and geographic locations. Protozoa, parasite checklist, host checklist, bibliography, Australia. Peter J. O’Donoghue, Department of Microbiology and Parasitology, The University of Queensland, St Lucia 4072, Australia; Robert D. Adlard, Protozoa Section, Queensland Museum, PO Box 3300, South Brisbane 4101, Australia; 31 January 2000. CONTENTS the literature for reports relevant to contemporary studies. Such problems could be avoided if all previous HOST-PARASITE CHECKLIST 5 records were consolidated into a single database. Most Mammals 5 researchers currently avail themselves of various Reptiles 21 electronic database and abstracting services but none Amphibians 26 include literature published earlier than 1985 and not all Birds 34 journal titles are covered in their databases. Fish 44 Invertebrates 54 Several catalogues of parasites in Australian PARASITE-HOST CHECKLIST 63 hosts have previously been published. -

Mitosomes in Entamoeba Histolytica Contain a Sulfate Activation Pathway
Mitosomes in Entamoeba histolytica contain a sulfate activation pathway Fumika Mi-ichi, Mohammad Abu Yousuf, Kumiko Nakada-Tsukui, and Tomoyoshi Nozaki1 Department of Parasitology, National Institute of Infectious Diseases, Tokyo 162-8640, Japan Edited by Andrew Roger, Dalhousie University, Halifax, NS, Canada, and accepted by the Editorial Board October 19, 2009 (received for review June 25, 2009) Hydrogenosomes and mitosomes are mitochondrion-related or- and M. balamuthi lack the ISC system, and instead possess the ganelles in anaerobic/microaerophilic eukaryotes with highly re- nitrogen fixation (NIF) system, which is most likely derived from an duced and divergent functions. The full diversity of their content ancestral nitrogen fixing -proteobacterium by lateral gene transfer and function, however, has not been fully determined. To under- (22, 24). Only 5 proteins have been demonstrated in E. histolytica stand the central role of mitosomes in Entamoeba histolytica,a mitosomes: Cpn60 (8–10, 12), Cpn10 (13), mitochondrial Hsp70 parasitic protozoon that causes amoebic dysentery and liver ab- (11, 15), pyridine nucleotide transhydrogenase (PNT) (2, 8), and scesses, we examined the proteomic profile of purified mitosomes. mitochondria carrier family (MCF, ADP/ATP transporter) (14), Using 2 discontinuous Percoll gradient centrifugation and MS and the central role of mitosomes in E. histolytica remains unknown. analysis, we identified 95 putative mitosomal proteins. Immuno- Analysis of the genome of E. histolytica has not revealed any fluorescence assay showed that 3 proteins involved in sulfate additional information regarding the function of mitosomes and activation, ATP sulfurylase, APS kinase, and inorganic pyrophos- thus, a proteomic analysis of mitosomes seems to be the best phatase, as well as sodium/sulfate symporter, involved in sulfate approach to understand its structure and function (1, 2). -

BMC Evolutionary Biology Biomed Central
BMC Evolutionary Biology BioMed Central Research article Open Access Complex distribution of EFL and EF-1α proteins in the green algal lineage Geoffrey P Noble, Matthew B Rogers and Patrick J Keeling* Address: Canadian Institute for Advanced Research, Department of Botany, University of British Columbia, 3529-6270 University Boulevard, Vancouver, BC V6T 1Z4, Canada Email: Geoffrey P Noble - [email protected]; Matthew B Rogers - [email protected]; Patrick J Keeling* - [email protected] * Corresponding author Published: 23 May 2007 Received: 5 February 2007 Accepted: 23 May 2007 BMC Evolutionary Biology 2007, 7:82 doi:10.1186/1471-2148-7-82 This article is available from: http://www.biomedcentral.com/1471-2148/7/82 © 2007 Noble et al; licensee BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Abstract Background: EFL (or elongation factor-like) is a member of the translation superfamily of GTPase proteins. It is restricted to eukaryotes, where it is found in a punctate distribution that is almost mutually exclusive with elongation factor-1 alpha (EF-1α). EF-1α is a core translation factor previously thought to be essential in eukaryotes, so its relationship to EFL has prompted the suggestion that EFL has spread by horizontal or lateral gene transfer (HGT or LGT) and replaced EF-1α multiple times. Among green algae, trebouxiophyceans and chlorophyceans have EFL, but the ulvophycean Acetabularia and the sister group to green algae, land plants, have EF-1α.