Harnessing Astrocytes and Müller Glial Cells in the Retina for Survival and Regeneration of Retinal Ganglion Cells
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Neuroinflammation Triggered by Β-Glucan/Dectin-1 Signaling Enables CNS Axon Regeneration
Neuroinflammation triggered by β-glucan/dectin-1 signaling enables CNS axon regeneration Katherine T. Baldwina,b,1, Kevin S. Carbajalc,d,1, Benjamin M. Segalc,d,2, and Roman J. Gigera,b,c,d,2 aDepartment of Cell and Developmental Biology, bCellular and Molecular Biology Graduate Program, cNeuroscience Graduate Program, and dHoltom-Garrett Program in Neuroimmunology, Department of Neurology, University of Michigan School of Medicine, Ann Arbor, MI 48109 Edited by Ben A. Barres, Stanford University School of Medicine, Stanford, CA, and approved January 22, 2015 (received for review December 22, 2014) Innate immunity can facilitate nervous system regeneration, yet growth can be undermined by concurrent toxicity (9). A deeper the underlying cellular and molecular mechanisms are not well understanding of these opposing effects will be important for understood. Here we show that intraocular injection of lipopoly- exploiting immunomodulatory pathways to promote neural re- saccharide (LPS), a bacterial cell wall component, or the fungal cell pair while minimizing bystander damage. wall extract zymosan both lead to rapid and comparable intravi- In the present study, we investigated the pathways that drive treal accumulation of blood-derived myeloid cells. However, when innate immune-mediated axon regeneration after ONC. We in- combined with retro-orbital optic nerve crush injury, lengthy duced sterile inflammation in the vitreous on the day of injury by growth of severed retinal ganglion cell (RGC) axons occurs only i.o. administration of zymosan or constituents of zymosan clas- in zymosan-injected mice, and not in LPS-injected mice. In mice sified as pathogen-associated molecular patterns (PAMPs). PAMPs dectin-1 deficient for the pattern recognition receptor but not are highly conserved microbial structures that serve as ligands for TLR2 , Toll-like receptor-2 ( ) zymosan-mediated RGC regeneration pattern recognition receptors (PRRs). -

Specification of Optic Nerve Oligodendrocyte Precursors by Retinal Ganglion Cell Axons
The Journal of Neuroscience, July 19, 2006 • 26(29):7619–7628 • 7619 Cellular/Molecular Specification of Optic Nerve Oligodendrocyte Precursors by Retinal Ganglion Cell Axons Limin Gao and Robert H. Miller Department of Neurosciences, Case Western Reserve University, Cleveland, Ohio 44106 Cell fate commitment in the developing CNS frequently depends on localized cell–cell interactions. In the avian visual system the optic nerve oligodendrocytes are derived from founder cells located at the floor of the third ventricle. Here we show that the induction of these founder cells is directly dependent on signaling from the retinal ganglion cell (RGC) axons. The appearance of oligodendrocyte precursor cells (OPCs) correlates with the projection of RGC axons, and early eye removal dramatically reduces the number of OPCs. In vitro signaling from retinal neurites induces OPCs in responsive tissue. Retinal axon induction of OPCs is dependent on sonic hedgehog (Shh) andneuregulinsignaling,andtheinhibitionofeithersignalreducesOPCinductioninvivoandinvitro.ThedependenceofOPCsonretinal axonal cues appears to be a common phenomenon, because ocular retardation (orJ) mice lacking optic nerve have dramatically reduced OPCs in the midline of the third ventricle. Key words: oligodendrocyte precursors; optic nerve; axon induction; sonic hedgehog; neuregulin; retinal ganglion cells Introduction contributes to the specification of ventral midline cells (Dale et al., During development the oligodendrocytes, the myelinating cells 1997); however, OPCs arise later -

Early Neuronal Processes Interact with Glia to Establish a Scaffold For
bioRxiv preprint doi: https://doi.org/10.1101/754416; this version posted August 31, 2019. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. Early neuronal processes interact with glia to establish a scaffold for orderly innervation of the cochlea Running Title: Glia and early cochlear wiring N. R. Druckenbrod1, E. B. Hale, O. O. Olukoya, W. E. Shatzer, and L.V. Goodrich* Department of Neurobiology, Harvard Medical School, Boston, MA, 02115 1Current address: Decibel Therapeutics, Boston, Ma, 02215 *correspondence should be addressed to [email protected] Keywords: neuron-glia interactions, axon guidance, spiral ganglion neuron, cochlea Authors’ contributions: This study was conceived of and designed by NRD and LVG. NRD, EBH, OOO, and WES performed experiments and analyzed results. LVG analyzed results and wrote the manuscript. bioRxiv preprint doi: https://doi.org/10.1101/754416; this version posted August 31, 2019. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. Summary: Although the basic principles of axon guidance are well established, it remains unclear how axons navigate with high fidelity through the complex cellular terrains that are encountered in vivo. To learn more about the cellular strategies underlying axon guidance in vivo, we analyzed the developing cochlea, where spiral ganglion neurons extend processes through a heterogeneous cellular environment to form tonotopically ordered connections with hair cells. -

Oligodendrocytes in Development, Myelin Generation and Beyond
cells Review Oligodendrocytes in Development, Myelin Generation and Beyond Sarah Kuhn y, Laura Gritti y, Daniel Crooks and Yvonne Dombrowski * Wellcome-Wolfson Institute for Experimental Medicine, Queen’s University Belfast, Belfast BT9 7BL, UK; [email protected] (S.K.); [email protected] (L.G.); [email protected] (D.C.) * Correspondence: [email protected]; Tel.: +0044-28-9097-6127 These authors contributed equally. y Received: 15 October 2019; Accepted: 7 November 2019; Published: 12 November 2019 Abstract: Oligodendrocytes are the myelinating cells of the central nervous system (CNS) that are generated from oligodendrocyte progenitor cells (OPC). OPC are distributed throughout the CNS and represent a pool of migratory and proliferative adult progenitor cells that can differentiate into oligodendrocytes. The central function of oligodendrocytes is to generate myelin, which is an extended membrane from the cell that wraps tightly around axons. Due to this energy consuming process and the associated high metabolic turnover oligodendrocytes are vulnerable to cytotoxic and excitotoxic factors. Oligodendrocyte pathology is therefore evident in a range of disorders including multiple sclerosis, schizophrenia and Alzheimer’s disease. Deceased oligodendrocytes can be replenished from the adult OPC pool and lost myelin can be regenerated during remyelination, which can prevent axonal degeneration and can restore function. Cell population studies have recently identified novel immunomodulatory functions of oligodendrocytes, the implications of which, e.g., for diseases with primary oligodendrocyte pathology, are not yet clear. Here, we review the journey of oligodendrocytes from the embryonic stage to their role in homeostasis and their fate in disease. We will also discuss the most common models used to study oligodendrocytes and describe newly discovered functions of oligodendrocytes. -

Chapter 8 Nervous System
Chapter 8 Nervous System I. Functions A. Sensory Input – stimuli interpreted as touch, taste, temperature, smell, sound, blood pressure, and body position. B. Integration – CNS processes sensory input and initiates responses categorizing into immediate response, memory, or ignore C. Homeostasis – maintains through sensory input and integration by stimulating or inhibiting other systems D. Mental Activity – consciousness, memory, thinking E. Control of Muscles & Glands – controls skeletal muscle and helps control/regulate smooth muscle, cardiac muscle, and glands II. Divisions of the Nervous system – 2 anatomical/main divisions A. CNS (Central Nervous System) – consists of the brain and spinal cord B. PNS (Peripheral Nervous System) – consists of ganglia and nerves outside the brain and spinal cord – has 2 subdivisions 1. Sensory Division (Afferent) – conducts action potentials from PNS toward the CNS (by way of the sensory neurons) for evaluation 2. Motor Division (Efferent) – conducts action potentials from CNS toward the PNS (by way of the motor neurons) creating a response from an effector organ – has 2 subdivisions a. Somatic Motor System – controls skeletal muscle only b. Autonomic System – controls/effects smooth muscle, cardiac muscle, and glands – 2 branches • Sympathetic – accelerator “fight or flight” • Parasympathetic – brake “resting and digesting” * 4 Types of Effector Organs: skeletal muscle, smooth muscle, cardiac muscle, and glands. III. Cells of the Nervous System A. Neurons – receive stimuli and transmit action potentials -

Fibrotic Scar in Neurodegenerative Diseases
MINI REVIEW published: 14 August 2020 doi: 10.3389/fimmu.2020.01394 Fibrotic Scar in Neurodegenerative Diseases Nadia D’Ambrosi* and Savina Apolloni* Department of Biology, Tor Vergata University, Rome, Italy The process of uncontrolled internal scarring, called fibrosis, is now emerging as a pathological feature shared by both peripheral and central nervous system diseases. In the CNS, damaged neurons are not replaced by tissue regeneration, and scar-forming cells such as endothelial cells, inflammatory immune cells, stromal fibroblasts, and astrocytes can persist chronically in brain and spinal cord lesions. Although this process was extensively described in acute CNS damages, novel evidence indicates the involvement of a fibrotic reaction in chronic CNS injuries as those occurring during neurodegenerative diseases, where inflammation and fibrosis fuel degeneration. In this mini review, we discuss recent advances around the role of fibrotic scar formation and function in different neurodegenerative conditions, particularly focusing on the rising role of scarring in the pathogenesis of amyotrophic lateral sclerosis, multiple sclerosis, and Alzheimer’s disease and highlighting the therapeutic relevance of targeting fibrotic scarring to slow and reverse neurodegeneration. Edited by: Keywords: Alzheimer’s disease, amyotrophic lateral sclerosis, astrocytes, fibroblasts, microglia, multiple sclerosis Anna-Maria Hoffmann-Vold, Oslo University Hospital, Norway Reviewed by: INTRODUCTION Carlo Chizzolini, Université de Genève, Switzerland Fibrosis identifies -

Astrogliosis in an Experimental Model of Hypovitaminosis B12: a Cellular Basis of Neurological Disorders Due to Cobalamin Deficiency
cells Article Astrogliosis in an Experimental Model of Hypovitaminosis B12: A Cellular Basis of Neurological Disorders Due to Cobalamin Deficiency Zuzanna Rzepka 1 , Jakub Rok 1 , Justyna Kowalska 1, Klaudia Banach 1, Justyna Magdalena Hermanowicz 2 , Artur Beberok 1 , Beata Sieklucka 2 , Dorota Gryko 3 and Dorota Wrze´sniok 1,* 1 Department of Pharmaceutical Chemistry, Faculty of Pharmaceutical Sciences in Sosnowiec, Medical University of Silesia in Katowice, Jagiello´nska4, 41-200 Sosnowiec, Poland; [email protected] (Z.R.); [email protected] (J.R.); [email protected] (J.K.); [email protected] (K.B.); [email protected] (A.B.) 2 Department of Pharmacodynamics, Medical University of Bialystok, Mickiewicza 2C, 15-222 Bialystok, Poland; [email protected] (J.M.H.); [email protected] (B.S.) 3 Institute of Organic Chemistry, Polish Academy of Science, Kasprzaka 44/52, 01-224 Warsaw, Poland; [email protected] * Correspondence: [email protected]; Tel.: +48-3-2364-1050 Received: 16 September 2020; Accepted: 7 October 2020; Published: 9 October 2020 Abstract: Cobalamin deficiency affects human physiology with sequelae ranging from mild fatigue to severe neuropsychiatric abnormalities. The cellular and molecular aspects of the nervous system disorders associated with hypovitaminosis B12 remain largely unknown. Growing evidence indicates that astrogliosis is an underlying component of a wide range of neuropathologies. Previously, we developed an in vitro model of cobalamin deficiency in normal human astrocytes (NHA) by culturing the cells with c-lactam of hydroxycobalamin (c-lactam OH-Cbl). We revealed a non-apoptotic activation of caspases (3/7, 8, 9) in cobalamin-deficient NHA, which may suggest astrogliosis. -
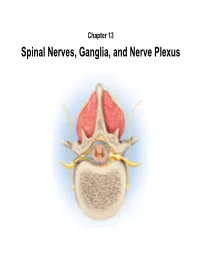
Spinal Nerves, Ganglia, and Nerve Plexus Spinal Nerves
Chapter 13 Spinal Nerves, Ganglia, and Nerve Plexus Spinal Nerves Posterior Spinous process of vertebra Posterior root Deep muscles of back Posterior ramus Spinal cord Transverse process of vertebra Posterior root ganglion Spinal nerve Anterior ramus Meningeal branch Communicating rami Anterior root Vertebral body Sympathetic ganglion Anterior General Anatomy of Nerves and Ganglia • Spinal cord communicates with the rest of the body by way of spinal nerves • nerve = a cordlike organ composed of numerous nerve fibers (axons) bound together by connective tissue – mixed nerves contain both afferent (sensory) and efferent (motor) fibers – composed of thousands of fibers carrying currents in opposite directions Anatomy of a Nerve Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Epineurium Perineurium Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Endoneurium Nerve Rootlets fiber Posterior root Fascicle Posterior root ganglion Anterior Blood root vessels Spinal nerve (b) Copyright by R.G. Kessel and R.H. Kardon, Tissues and Organs: A Text-Atlas of Scanning Electron Microscopy, 1979, W.H. Freeman, All rights reserved Blood vessels Fascicle Epineurium Perineurium Unmyelinated nerve fibers Myelinated nerve fibers (a) Endoneurium Myelin General Anatomy of Nerves and Ganglia • nerves of peripheral nervous system are ensheathed in Schwann cells – forms neurilemma and often a myelin sheath around the axon – external to neurilemma, each fiber is surrounded by -

Spinal Cord Organization
Lecture 4 Spinal Cord Organization The spinal cord . Afferent tract • connects with spinal nerves, through afferent BRAIN neuron & efferent axons in spinal roots; reflex receptor interneuron • communicates with the brain, by means of cell ascending and descending pathways that body form tracts in spinal white matter; and white matter muscle • gives rise to spinal reflexes, pre-determined gray matter Efferent neuron by interneuronal circuits. Spinal Cord Section Gross anatomy of the spinal cord: The spinal cord is a cylinder of CNS. The spinal cord exhibits subtle cervical and lumbar (lumbosacral) enlargements produced by extra neurons in segments that innervate limbs. The region of spinal cord caudal to the lumbar enlargement is conus medullaris. Caudal to this, a terminal filament of (nonfunctional) glial tissue extends into the tail. terminal filament lumbar enlargement conus medullaris cervical enlargement A spinal cord segment = a portion of spinal cord that spinal ganglion gives rise to a pair (right & left) of spinal nerves. Each spinal dorsal nerve is attached to the spinal cord by means of dorsal and spinal ventral roots composed of rootlets. Spinal segments, spinal root (rootlets) nerve roots, and spinal nerves are all identified numerically by th region, e.g., 6 cervical (C6) spinal segment. ventral Sacral and caudal spinal roots (surrounding the conus root medullaris and terminal filament and streaming caudally to (rootlets) reach corresponding intervertebral foramina) collectively constitute the cauda equina. Both the spinal cord (CNS) and spinal roots (PNS) are enveloped by meninges within the vertebral canal. Spinal nerves (which are formed in intervertebral foramina) are covered by connective tissue (epineurium, perineurium, & endoneurium) rather than meninges. -

Neuron Morphology Influences Axon Initial Segment Plasticity1,2,3
New Research Neuronal Excitability Neuron Morphology Influences Axon Initial Segment Plasticity1,2,3 Allan T. Gulledge1 and Jaime J. Bravo2 DOI:http://dx.doi.org/10.1523/ENEURO.0085-15.2016 1Department of Physiology and Neurobiology, Geisel School of Medicine at Dartmouth, Dartmouth-Hitchcock Medical Center, Lebanon, New Hampshire 03756, and 2Thayer School of Engineering at Dartmouth, Hanover, New Hampshire 03755 Visual Abstract In most vertebrate neurons, action potentials are initiated in the axon initial segment (AIS), a specialized region of the axon containing a high density of voltage-gated sodium and potassium channels. It has recently been proposed that neurons use plasticity of AIS length and/or location to regulate their intrinsic excitability. Here we quantify the impact of neuron morphology on AIS plasticity using computational models of simplified and realistic somatodendritic morphologies. In small neurons (e.g., den- tate granule neurons), excitability was highest when the AIS was of intermediate length and located adjacent to the soma. Conversely, neu- rons having larger dendritic trees (e.g., pyramidal neurons) were most excitable when the AIS was longer and/or located away from the soma. For any given somatodendritic morphology, increasing dendritic mem- brane capacitance and/or conductance favored a longer and more distally located AIS. Overall, changes to AIS length, with corresponding changes in total sodium conductance, were far more effective in regulating neuron excitability than were changes in AIS location, while dendritic capacitance had a larger impact on AIS performance than did dendritic conductance. The somatodendritic influence on AIS performance reflects modest soma- to-AIS voltage attenuation combined with neuron size-dependent changes in AIS input resistance, effective membrane time constant, and isolation from somatodendritic capacitance. -

Digital Neuroanatomy
DIGITAL NEUROANATOMY LM NEUROHISTOLOGY George R. Leichnetz, Ph.D. Professor, Department of Anatomy & Neurobiology Virginia Commonwealth University 2004 Press the Å and Æ keys on your keyboard to navigate through this lecture There are three morphological types of neurons in the nervous system: unipolar, bipolar, and multipolar. While all three types can be found in the PNS, the CNS only contains multipolar neurons. CNS multipolar neurons vary greatly in morphology: eg. spinal cord motor neurons, pyramidal cells of cerebral cortex, Purkinje cells of cerebellar cortex. The supportive cells of the CNS are neuroglia: astrocytes, oligodendrocytes, and microglia. Oligodendrocytes myelinate CNS axons, while Schwann cells myelinate PNS axons. Protoplasmic astrocytes predominate in CNS gray matter, while fibrous astrocytes predominate in CNS white matter. The ventral horn of the spinal cord Lumbar Spinal Cord contains multipolar motor neurons. Dorsal White Horn Matter Gray Matter Ventral Kluver- Horn Barrera Stain (LFB & CV) Cell bodies of motor neurons Dendrite Gray Matter Dendrite Perineuronal H & E oligodendrocyte Stain Nucleus, nucleolus Abundant Nissl substance Dorsal root Dorsal root ganglion Ventral root Gray Matter: contains White Matter: cell bodies of neurons contains x-sections of myelinated axons Multipolar motor neurons Silver Stain of the ventral horn Large multipolar motor neurons of the spinal cord ventral horn: see nucleus, nucleolus, abundant Nissl substance, multiple tapering processes (dendrites). The axon hillock (origin of axon from cell body) lacks Nissl. Nucleus Axon hillock dendrites Nucleolus Nissl substance (RER) Kluver-Barrera Stain (Luxol Fast Blue and Cresyl Violet) Thoracic Spinal Cord The nucleus dorsalis of Clarke, located in the base of the dorsal horn of the thoracic spinal cord, contains multipolar neurons whose nucleus is eccentric and Nissl around the periphery of the cell body (as if it were chromatolytic). -

Reversing Glial Scar Back to Neural Tissue Through Neurod1-Mediated Astrocyte-To-Neuron Conversion
bioRxiv preprint doi: https://doi.org/10.1101/261438; this version posted February 7, 2018. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. Reversing Glial Scar Back To Neural Tissue Through NeuroD1-Mediated Astrocyte-To-Neuron Conversion Lei Zhang1, Zhuofan Lei1, Ziyuan Guo1, Zifei Pei1, Yuchen Chen1, Fengyu Zhang1, Alice Cai1, Yung Kin Mok1, Grace Lee1, Vishal Swaminnathan1, Fan Wang1, Yuting Bai1, Gong Chen1,* 1. Department of Biology, Huck Institute of Life Sciences, Pennsylvania State University, University Park, PA 16802, USA. Keywords: glial scar, NeuroD1, stab injury, in vivo conversion, glia to neuron ratio, brain repair * Correspondence should be addressed to: Gong Chen, Ph.D. Professor and Verne M. Willaman Chair in Life Sciences Department of Biology, Huck Institute of Life Sciences, The Pennsylvania State University, University Park, PA 16802, USA. Email: [email protected] Phone: 814-865-2488 Website: http://bio.psu.edu/directory/guc2 1 bioRxiv preprint doi: https://doi.org/10.1101/261438; this version posted February 7, 2018. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. ABSTRACT Nerve injury often causes neuronal loss and glial proliferation, disrupting the delicate balance between neurons and glial cells in the brain. Recently, we have developed an innovative technology to convert internal reactive glial cells into functional neurons inside the mouse brain. Here, we further demonstrate that such glia-to-neuron conversion can rebalance neuron-glia ratio and reverse glial scar back to neural tissue.