Dry Tortugas National Parks
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Socan Monitoring Workshop
SOCAN MONITORING WORKSHOP Image credit: Lauren Valentino, NOAA/AOML 2/28/17 WorksHop Report SOCAN MONITORING WORKSHOP SOCAN MONITORING WORKSHOP IDENTIFYING PRIORITY LOCATIONS FOR OCEAN ACIDIFICATION MONITORING IN THE U.S. SOUTHEAST SUMMARY The Southeast Ocean and Coastal Acidification Network (SOCAN) held a worksHop in CHarleston, SoutH Carolina to facilitate discussion on priority locations for ocean acidification monitoring in tHe SoutHeast. The discussion included identification of key gradients in physical, chemical and biological parameters along tHe SoutHeast coast, a review of current monitoring efforts, and an assessment of stakeHolder needs. Sixteen monitoring locations were identified as potential acidification monitoring locations (see page 12). The following three monitoring locations were HigHligHted as priority sites tHat would furtHer our understanding of the chemistry and regional drivers of ocean acidification and address stakeholder needs: (1) Sapelo Island, GA (2) Gulf Stream, offsHore of Gray’s Reef, GA (3) Biscayne National Park, FL The workshop concluded witH a discussion of logistics and opportunities to pursue monitoring at tHe recommended locations. A copy of tHe agenda is included in Appendix 1. PROCEEDINGS Approximately 16 experts gatHered for tHe SOCAN Monitoring Workshop to outline recommendations for priority ocean acidification monitoring locations in tHe SoutHeast (Attendee List, Appendix 2). The worksHop began witH introductory remarks regarding tHe structure and responsibilities of SOCAN and SECOORA. Following the introductory remarks, participants reviewed the proposed agenda; no modifications were made. The first half-day was spent reviewing tHe state of ocean acidification science and regional response. Kim Yates sHared a syntHesis of tHe 2016 SOCAN State of the Science meeting, wHicH included a review of webinars and key findings related to OA chemistry, modeling and organismal response. -
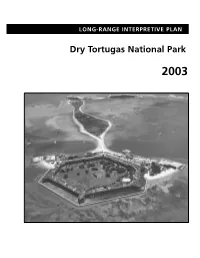
Long-Range Interpretive Plan, Dry Tortugas National Park
LONG-RANGE INTERPRETIVE PLAN Dry Tortugas National Park 2003 Cover Photograph: Aerial view of Fort Jefferson on Garden Key (fore- ground) and Bush Key (background). COMPREHENSIVE INTERPRETIVE PLAN Dry Tortugas National Park 2003 LONG-RANGE INTERPRETIVE PLAN Dry Tortugas National Park 2003 Prepared by: Department of Interpretive Planning Harpers Ferry Design Center and the Interpretive Staff of Dry Tortugas National Park and Everglades National Park INTRODUCTION About 70 miles west of Key West, Florida, lies a string of seven islands called the Dry Tortugas. These sand and coral reef islands, or keys, along with 100 square miles of shallow waters and shoals that surround them, make up Dry Tortugas National Park. Here, clear views of water and sky extend to the horizon, broken only by an occasional island. Below and above the horizon line are natural and historical treasures that continue to beckon and amaze those visitors who venture here. Warm, clear, shallow, and well-lit waters around these tropical islands provide ideal conditions for coral reefs. Tiny, primitive animals called polyps live in colonies under these waters and form skeletons from cal- cium carbonate which, over centuries, create coral reefs. These reef ecosystems support a wealth of marine life such as sea anemones, sea fans, lobsters, and many other animal and plant species. Throughout these fragile habitats, colorful fishes swim, feed, court, and thrive. Sea turtles−−once so numerous they inspired Spanish explorer Ponce de León to name these islands “Las Tortugas” in 1513−−still live in these waters. Loggerhead and Green sea turtles crawl onto sand beaches here to lay hundreds of eggs. -

Dry Tortugas U.S
National Park Service Dry Tortugas U.S. Department of the Interior Dry Tortugas National Park P.O. Box 6208 Key West, FL 33041 Park Regulations Welcome to the Park Welcome to Dry Tortugas National Park! This is a special place, and requires care from all who visit. The following is a summary of those regulations most important to park visitors. These regulations are necessary for protecting the fragile natural and historical features within Dry Tortugas National Park, and for ensuring your safety. When in doubt, ask park staff for additional information. Have a safe and enjoyable visit! Public Use Areas and Fort Jefferson and Garden Key Bush Key Closures The fort interior is open sunrise to sunset. Pets, food, Closed January 16-October 14 for bird nesting. and drinks, are not permitted inside the fort. Service and residential areas are closed to the public. The Hospital & Long Keys moat is closed to all entries and activities. The Special Closed year round. Visitors should remain 100 feet Protection Zones for shark breeding and corals on off shore of all closed islands. the east and southeast side of Garden Key harbor are closed to all vessels. Loggerhead Key Open year round during daylight hours only. Middle and East Key Dock and all structures are closed to public use. Closed April 1- October 15 for turtle nesting. Exploring on foot is limited to developed trails and shoreline between water and high tide line only. Fishing Important Reminders Park Areas Closed to Fishing All State of Florida saltwater fishing laws and regula- All fishing is prohibited within the Research Natural tions apply except as modified below. -

Sample Itinerary: Dry Tortugas – One Week*
Sample Itinerary: Dry Tortugas – One Week* Day 1 – Depart out of Marathon in the evening and sail to the Dry Tortugas. (107 NM) It will take anywhere from 12 to 16 hours to make this passage. You should try to arrive in day light hours so that your approach is easier. Day 2 – Arrive at the island group known as the Dry Tortugas. This is the most isolated and least visited national park in the United States. You should plan to spend two nights here but if the weather deteriorates be flexible enough to cut it short and head back to Key West. There are no services out on these islands so you should plan to be self sufficient while in this remote area. Day 3 – Go ashore on Garden Key and tour Fort Jefferson National Monument. This was originally built in the mid 1800’s as a military fort to be used by Union forces during the Civil War. The fort was converted to a prison whose most famous inmate was Dr. Mudd of Lincoln’s time. Day 4 – Snorkel the reefs surrounding the islands. Go ashore on Loggerhead Key and/or Hospital Key for a cookout, just be sure to clean up well and leave as little of a footprint as possible. Bush Key offers great bird watching from your yacht. Bush Key is a bird sanctuary and landing is prohibited. Day 5 – Depart the Dry Tortugas early in the morning and sail to the uninhabited island group of the Marquesas. (44 NM) Be careful of Rebecca Shoal on the way and arrive before dark. -

Central Keys Area Reasonable Assurance Documentation
Central Keys Area Reasonable Assurance Documentation F K R A D FKRAD Program December 2008 Prepared for FLORIDA DEPARTMENT OF ENVIRONMENTAL PROTECTION Watershed Management Bureau Tallahassee, Florida Prepared by Camp, Dresser & McKee, Inc. URS Corporation Southern 1715 N. Westshore Boulevard 7650 Courtney Campbell Causeway Tampa, Florida 33607 Tampa, Florida 33607 Florida Department of Environmental Protection REASONABLE ASSURANCE DOCUMENTATION CENTRAL KEYS AREA F K R A D December 2008 ACKNOWLEDGEMENTS The Central Keys Area Reasonable Assurance Document was developed under the direction of Mr. Fred Calder and Mr. Pat Fricano of the Florida Department of Environmental Protection (FDEP) with the assistance of the following stakeholder’s representatives: Ms. Suzy Thomas, Director of Community Services and Mr. Mike Hatfield, P.E., Weiler Engineering, City of Marathon Mr. Clyde Burnett, Mayor/City Administrator, City of Key Colony Beach; Mr. Philip “Skip” Haring, Assistant to Mayor City of Layton; Ms. Elizabeth Wood, P.E., Wastewater Section Chief, Monroe County; Mr. Jaime Barrera, FDOT District VI; and Mr. Fred Hand, Bureau of Facilities, FDEP. The authors gratefully acknowledge the comments review comments of the Stakeholders technical representatives and Mr. Gus Rios, Manager of FDEP’s Marathon Service Office. The document was developed a collaborative effort led by Mr. Scott McClelland of CDM Inc. and Mr. Stephen Lienhart of URS Corporation. S:\FDEP\Central RAD\FKRAD Central Cover and Prelim.doc i Florida Department of Environmental Protection REASONABLE ASSURANCE DOCUMENTATION CENTRAL KEYS AREA F K R A D December 2008 Central Keys Area Stakeholders Documents As a measure of reasonable assurance and support of this document, the stakeholders in the Central Keys Area (City of Marathon, City of Key Colony Beach, City of Layton, Monroe County, FDOT and the Florida State Parks Service) have provided signed documents confirming that the management activities identified in this document indeed reflect the commitments of the stakeholders. -

Collier Miami-Dade Palm Beach Hendry Broward Glades St
Florida Fish and Wildlife Conservation Commission F L O R ID A 'S T U R N P IK E er iv R ee m Lakewood Park m !( si is O K L D INDRIO ROAD INDRIO RD D H I N COUNTY BCHS Y X I L A I E O W L H H O W G Y R I D H UCIE BLVD ST L / S FT PRCE ILT SRA N [h G Fort Pierce Inlet E 4 F N [h I 8 F AVE "Q" [h [h A K A V R PELICAN YACHT CLUB D E . FORT PIERCE CITY MARINA [h NGE AVE . OKEECHOBEE RA D O KISSIMMEE RIVER PUA NE 224 ST / CR 68 D R !( A D Fort Pierce E RD. OS O H PIC R V R T I L A N N A M T E W S H N T A E 3 O 9 K C A R-6 A 8 O / 1 N K 0 N C 6 W C W R 6 - HICKORY HAMMOCK WMA - K O R S 1 R L S 6 R N A E 0 E Lake T B P U Y H D A K D R is R /NW 160TH E si 68 ST. O m R H C A me MIDWAY RD. e D Ri Jernigans Pond Palm Lake FMA ver HUTCHINSON ISL . O VE S A t C . T I IA EASY S N E N L I u D A N.E. 120 ST G c I N R i A I e D South N U R V R S R iv I 9 I V 8 FLOR e V ESTA DR r E ST. -

Federal Register/Vol. 63, No. 158/Monday, August 17, 1998
43870 Federal Register / Vol. 63, No. 158 / Monday, August 17, 1998 / Rules and Regulations U.S.C. 553) because: (1) The 1998±99 SUMMARY: The National Oceanic and effective July 1, 1997, and codified at 15 fiscal year began on July 1, 1998, and Atmospheric Administration amends CFR Part 922, Subpart P. the marketing order requires that the the regulations for the Florida Keys In September 1997, NOAA became rate of assessment for each fiscal year National Marine Sanctuary (FKNMS or aware that significant injury to, and apply to all assessable papayas handled Sanctuary) to reinstate and make destruction of, living coral on the during such fiscal year; (2) the permanent the temporary prohibition on Tortugas Bank, west of the Dry Tortugas Committee needs to have sufficient anchoring by vessels 50 meters or National Park, was being caused by the funds to pay its expenses which are greater in registered length on Tortugas anchoring of vessels 50 meters or greater incurred on a continuous basis; and (3) Bank. The preamble to this rule contains in registered length. handlers are aware of this action which an environmental assessment for this Section 922.165 of the Sanctuary was recommended by the Committee at action. The intent of this rule is to regulations provides that, where a public meeting and is similar to other protect the coral reef at Tortugas Bank. necessary to prevent or minimize the assessment rate actions issued in past The proposed rule was published on destruction of, loss of, or injury to a years. February 11, 1998 and the comment Sanctuary resources, any and all List of Subjects in 7 CFR Part 928 period ended on March 13, 1998. -

Coast Guard, DHS § 7.100
Coast Guard, DHS § 7.100 the easternmost extremity of Black- (e) A line drawn across the seaward beard Island at Northeast Point. extremity of the Sebastian Inlet Jet- (d) A line drawn from the southern- ties. most extremity of Blackbeard Island to (f) A line drawn from the seaward ex- latitude 31°19.4′ N. longitude 81°11.5′ W. tremity of the Fort Pierce Inlet North (Doboy Sound Lighted Buoy ‘‘D’’); Jetty to latitude 27°28.5′ N. longitude thence to latitude 31°04.1′ N. longitude 80°16.2′ W. (Fort Pierce Inlet Lighted 81°16.7′ W. (St. Simons Lighted Whistle Whistle Buoy ‘‘2’’); thence to the tank Buoy ‘‘ST S’’). located in approximate position lati- tude 27°27.2′ N. longitude 80°17.2′ W. § 7.85 St. Simons Island, GA to Little (g) A line drawn from the seaward ex- Talbot Island, FL. tremity of St. Lucie Inlet north jetty (a) A line drawn from latitude 31°04.1′ to latitude 27°10′ N. longitude 80°08.4′ N. longitude 81°16.7′ W. (St. Simons W. (St. Lucie Inlet Entrance Lighted Lighted Whistle Buoy ‘‘ST S’’) to lati- Whistle Buoy ‘‘2’’); thence to Jupiter tude 30°42.7′ N. longitude 81°19.0′ W. (St. Island bearing approximately 180° true. Mary’s Entrance Lighted Whistle Buoy (h) A line drawn from the seaward ex- ‘‘1’’); thence to Amelia Island Light. tremity of Jupiter Inlet North Jetty to (b) A line drawn from the southern- the northeast extremity of the con- most extremity of Amelia Island to crete apron on the south side of Jupiter latitude 30°29.4′ N. -

Biscayne National Park's Fishery Management Plan
WATER Credit: iStock.com/Matt_Potenski Biscayne National Park’s Fishery 77% Management Plan OF 35 FISH STOCKS STUDIED Biscayne National Park is a national treasure 5 WITHIN THE PARK ened, some on the verge of collapse. If ac- and home to part of the third largest barrier tion isn’t taken and fish populations and ARE OVERFISHED, reef ecosystem in the world. 95% water, MOST CAPTURED habitat health continue to decline, the Park Biscayne is the largest marine park in the BEFORE THEY Service will have failed at its fundamental National Park System, created to protect, “a HAD A CHANCE TO mission to protect Biscayne National Park in SPAWN.1 rare combination of terrestrial, marine, and perpetuity for all Americans. Implementing amphibious life in a tropical setting of great a marine reserve and science-based fisheries natural beauty” for present and future management policies are key to protecting 3 generations. In addition to its ecological and restoring Biscayne’s marine resources. value, the park is a significant economic 95% driver, supporting a variety of economic and Biscayne’s Fishery Management Plan DECLINE IN REEF recreation activities, such as fishing, diving, Biscayne National Park’s Fishery Manage- FISH CATCHES IN snorkeling, and boating. According to a ment Plan (FMP), finalized back in 2014, BISCAYNE SINCE National Park Service report, in 2018 over aims to increase the average size and abun- THE 1960s.2 450,000 visitors to Biscayne spent more than dance of target fish species in the park by $30 million, supporting 398 local jobs and 20% by creating park-specific fisheries man- generating more than $42 million for the lo- agement regulations and reducing fishing- cal economy.4 related habitat degradation. -

Restoring Southern Florida's Native Plant Heritage
A publication of The Institute for Regional Conservation’s Restoring South Florida’s Native Plant Heritage program Copyright 2002 The Institute for Regional Conservation ISBN Number 0-9704997-0-5 Published by The Institute for Regional Conservation 22601 S.W. 152 Avenue Miami, Florida 33170 www.regionalconservation.org [email protected] Printed by River City Publishing a division of Titan Business Services 6277 Powers Avenue Jacksonville, Florida 32217 Cover photos by George D. Gann: Top: mahogany mistletoe (Phoradendron rubrum), a tropical species that grows only on Key Largo, and one of South Florida’s rarest species. Mahogany poachers and habitat loss in the 1970s brought this species to near extinction in South Florida. Bottom: fuzzywuzzy airplant (Tillandsia pruinosa), a tropical epiphyte that grows in several conservation areas in and around the Big Cypress Swamp. This and other rare epiphytes are threatened by poaching, hydrological change, and exotic pest plant invasions. Funding for Rare Plants of South Florida was provided by The Elizabeth Ordway Dunn Foundation, National Fish and Wildlife Foundation, and the Steve Arrowsmith Fund. Major funding for the Floristic Inventory of South Florida, the research program upon which this manual is based, was provided by the National Fish and Wildlife Foundation and the Steve Arrowsmith Fund. Nemastylis floridana Small Celestial Lily South Florida Status: Critically imperiled. One occurrence in five conservation areas (Dupuis Reserve, J.W. Corbett Wildlife Management Area, Loxahatchee Slough Natural Area, Royal Palm Beach Pines Natural Area, & Pal-Mar). Taxonomy: Monocotyledon; Iridaceae. Habit: Perennial terrestrial herb. Distribution: Endemic to Florida. Wunderlin (1998) reports it as occasional in Florida from Flagler County south to Broward County. -

Dry Tortugas U.S
National Park Service Dry Tortugas U.S. Department of the Interior Dry Tortugas National Park Life on Loggerhead Loggerhead Light The largest island in the Dry Tortugas, Loggerhead Key has seen many changes in its history. From a site of shipwrecks, to a world-class lighthouse installation, to a cutting-edge marine research laboratory, to a goal for Cuban refugees, Loggerhead Key is a part of the crossroads that are the Dry Tortugas. Loggerhead Lives Named for its abundance of loggerhead sea with a 2nd order bivalve lens in 1909), the new turtles, Loggerhead Key has long been a haven light was observed at a distance of 53 miles. The for wildlife. Migrating birds flock here on their 2nd order lens is now on display at the National way north, and coral fishes are abundant in the Aids to Navigation School in Yorktown, VA. coral reefs just offshore. But sea turtles are perhaps the most abundant species on the While the lighthouse was under construction, island, with approximately 250 nests yielding a contractor finished construction of a house 15,000 hatchlings each summer. The Dry Tortu- and kitchen (as seen in the 1870 image above). gas support the largest green and loggerhead sea The main house burned in 1945, but the kitchen turtle nesting grounds in the Florida Keys. (the smaller brick house in the image above) still stands and is occupied by National Park Service Mariners were often attracted to the sea turtles volunteer caretakers. A "new" lighthouse on Loggerhead as a food source for their sea keepers house to the north of the lighthouse voyage, but they found the Tortugas to be a was built for the keeper's family in the 1920s. -

Keys Sanctuary 25 Years of Marine Preservation National Parks Turn 100 Offbeat Keys Names Florida Keys Sunsets
Keys TravelerThe Magazine Keys Sanctuary 25 Years of Marine Preservation National Parks Turn 100 Offbeat Keys Names Florida Keys Sunsets fla-keys.com Decompresssing at Bahia Honda State Park near Big Pine Key in the Lower Florida Keys. ANDY NEWMAN MARIA NEWMAN Keys Traveler 12 The Magazine Editor Andy Newman Managing Editor 8 4 Carol Shaughnessy ROB O’NEAL ROB Copy Editor Buck Banks Writers Julie Botteri We do! Briana Ciraulo Chloe Lykes TIM GROLLIMUND “Keys Traveler” is published by the Monroe County Tourist Development Contents Council, the official visitor marketing agency for the Florida Keys & Key West. 4 Sanctuary Protects Keys Marine Resources Director 8 Outdoor Art Enriches the Florida Keys Harold Wheeler 9 Epic Keys: Kiteboarding and Wakeboarding Director of Sales Stacey Mitchell 10 That Florida Keys Sunset! Florida Keys & Key West 12 Keys National Parks Join Centennial Celebration Visitor Information www.fla-keys.com 14 Florida Bay is a Must-Do Angling Experience www.fla-keys.co.uk 16 Race Over Water During Key Largo Bridge Run www.fla-keys.de www.fla-keys.it 17 What’s in a Name? In Marathon, Plenty! www.fla-keys.ie 18 Visit Indian and Lignumvitae Keys Splash or Relax at Keys Beaches www.fla-keys.fr New Arts District Enlivens Key West ach of the Florida Keys’ regions, from Key Largo Bahia Honda State Park, located in the Lower Keys www.fla-keys.nl www.fla-keys.be Stroll Back in Time at Crane Point to Key West, features sandy beaches for relaxing, between MMs 36 and 37. The beaches of Bahia Honda Toll-Free in the U.S.