Cytokines As Mediators of Neuroinflammation in Experimental Autoimmune Encephalomyelitis
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

(CS-ⅣA-Be), a Novel IL-6R Antagonist, Inhibits IL-6/STAT3
Author Manuscript Published OnlineFirst on February 29, 2016; DOI: 10.1158/1535-7163.MCT-15-0551 Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Chikusetsusaponin Ⅳa butyl ester (CS-Ⅳa-Be), a novel IL-6R antagonist, inhibits IL-6/STAT3 signaling pathway and induces cancer cell apoptosis Jie Yang 1, 2, Shihui Qian 2, Xueting Cai 1, 2, Wuguang Lu 1, 2, Chunping Hu 1, 2, * Xiaoyan Sun1, 2, Yang Yang1, 2, Qiang Yu 3, S. Paul Gao 4, Peng Cao 1, 2 1. Affiliated Hospital of Integrated Traditional Chinese and Western Medicine, Nanjing University of Chinese Medicine, Nanjing 210028, China 2. Laboratory of Cellular and Molecular Biology, Jiangsu Province Academy of Traditional Chinese Medicine, Nanjing 210028, China 3. Shanghai Institute of Materia Medical, Chinese Academy of Sciences, Shanghai, 201203, China 4. Human Oncology and Pathogenesis Program, Memorial Sloan-Kettering Cancer Center, New York, NY10065, USA Running title: CS-Ⅳa-Be, a novel IL-6R antagonist, inhibits IL-6/STAT3 Keywords: Chikusetsusaponin Ⅳ a butyl ester (CS- Ⅳ a-Be), STAT3, IL-6R, antagonist, cancer Grant support: P. Cao received Jiangsu Province Funds for Distinguished Young Scientists (BK20140049) grant, J. Yang received National Natural Science Foundation of China (No. 81403151) grant, and X.Y. Sun received National Natural Science Foundation of China (No. 81202576) grant. Corresponding author: Peng Cao Institute: Laboratory of Cellular and Molecular Biology, Jiangsu Province Academy of Traditional Chinese Medicine, Nanjing 210028, Jiangsu, China Mailing address: 100#, Shizi Street, Hongshan Road, Nanjing, Jiangsu, China Tel: +86-25-85608666 Fax: +86-25-85608666 Email address: [email protected] The first co-authors: Jie Yang and Shihui Qian The authors disclose no potential conflicts of interest. -

Kcnorman 1.Pdf
A Computational Systems Approach to Elucidate New Mechanisms Involved in Progressive Lung Disease by Katy Norman A dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy (Biomedical Engineering) in the University of Michigan 2020 Doctoral Committee: Assistant Professor Kelly Arnold, Chair Professor Jeffrey Curtis Professor Jennifer Linderman Professor Bethany Moore Professor David Sept Katy C. Norman [email protected] ORCID iD: 0000-0001-8841-0212 © Katy C. Norman 2020 Acknowledgements When I look back on myself in 2015 as I just entered the PhD program, I have to give a little laugh about all the surprises and changes that have happened along the way. I have grown more than I could ever have imagined – I have become so much more comfortable and confident in my communication skills, in my scientific skills, and in myself. These changes and this growth could not have happened without my huge support network, and I am honored to have the chance to thank everyone for what they have given me and for what they have helped me achieve. I would first like to thank my adviser, Kelly Arnold, for giving me a chance to try out computational systems biology research when I had absolutely no background in the field coming in. I appreciate how you were easily able to create a space where I felt comfortable to learn, to ask questions, and to make mistakes as I worked towards gaining expertise in this field. Thank you for always being there for me with extra support when my nerves were getting the best of me before big presentations or before a big deadline, and for supporting my involvement in activities outside of the lab as well. -

GP130 Cytokines in Breast Cancer and Bone
cancers Review GP130 Cytokines in Breast Cancer and Bone Tolu Omokehinde 1,2 and Rachelle W. Johnson 1,2,3,* 1 Program in Cancer Biology, Vanderbilt University, Nashville, TN 37232, USA; [email protected] 2 Vanderbilt Center for Bone Biology, Department of Medicine, Division of Clinical Pharmacology, Vanderbilt University Medical Center, Nashville, TN 37232, USA 3 Department of Medicine, Division of Clinical Pharmacology, Vanderbilt University Medical Center, Nashville, TN 37232, USA * Correspondence: [email protected]; Tel.: +1-615-875-8965 Received: 14 December 2019; Accepted: 29 January 2020; Published: 31 January 2020 Abstract: Breast cancer cells have a high predilection for skeletal homing, where they may either induce osteolytic bone destruction or enter a latency period in which they remain quiescent. Breast cancer cells produce and encounter autocrine and paracrine cytokine signals in the bone microenvironment, which can influence their behavior in multiple ways. For example, these signals can promote the survival and dormancy of bone-disseminated cancer cells or stimulate proliferation. The interleukin-6 (IL-6) cytokine family, defined by its use of the glycoprotein 130 (gp130) co-receptor, includes interleukin-11 (IL-11), leukemia inhibitory factor (LIF), oncostatin M (OSM), ciliary neurotrophic factor (CNTF), and cardiotrophin-1 (CT-1), among others. These cytokines are known to have overlapping pleiotropic functions in different cell types and are important for cross-talk between bone-resident cells. IL-6 cytokines have also been implicated in the progression and metastasis of breast, prostate, lung, and cervical cancer, highlighting the importance of these cytokines in the tumor–bone microenvironment. This review will describe the role of these cytokines in skeletal remodeling and cancer progression both within and outside of the bone microenvironment. -

Due to Interleukin-6 Type Cytokine Redundancy Only Glycoprotein 130 Receptor Blockade Efficiently Inhibits Myeloma Growth
Plasma Cell Disorders SUPPLEMENTARY APPENDIX Due to interleukin-6 type cytokine redundancy only glycoprotein 130 receptor blockade efficiently inhibits myeloma growth Renate Burger, 1 Andreas Günther, 1 Katja Klausz, 1 Matthias Staudinger, 1 Matthias Peipp, 1 Eva Maria Murga Penas, 2 Stefan Rose-John, 3 John Wijdenes 4 and Martin Gramatzki 1 1Division of Stem Cell Transplantation and Immunotherapy, Department of Internal Medicine II, Christian-Albrechts-University Kiel and University Medical Center Schleswig-Holstein, Kiel, Germany; 2Institute of Human Genetics, Christian-Albrechts-University Kiel and Uni - versity Medical Center Schleswig-Holstein, Kiel, Germany; 3Department of Biochemistry, Christian-Albrechts-University of Kiel, Medical Faculty, Germany and 4Gen-Probe/Diaclone SAS, Besançon, France ©2017 Ferrata Storti Foundation. This is an open-access paper. doi:10.3324/haematol. 2016.145060 Received: February 25, 2016. Accepted: September 14, 2016. Pre-published: September 22, 2016. Correspondence: [email protected] SUPPLEMENTARY METHODS Cell lines and culture Cell lines INA-6, INA-6.Tu1 and B9 were cultivated in RPMI-1640 with GlutaMax™-I, 25 mM HEPES (Gibco®/Life Technologies GmbH, Darmstadt, Germany), 10% (v/v) heat-inactivated fetal bovine serum (FBS) (HyClone; Perbio Science, Erembodegen, Belgium), and antibiotics (R10+ medium) supplemented with 2.5 ng/ml recombinant huIL-6 (Gibco®/Life Technologies GmbH, Darmstadt, Germany). The cell lines are routinely confirmed to be negative for mycoplasma contamination (Venor™GeM Mycoplasma Detection Kit, Sigma-Aldrich, St. Louis, MO). Cytokines and other reagents Recombinant huIL-6 was purchased from Gibco®/Life Technologies (Darmstadt, Germany), huLIF was from Reliatech (Wolfenbüttel, Germany). Recombinant muIL-6 was obtained from Peprotech (Rocky Hill, NJ), and soluble muIL-6R was from R&D Systems (Minneapolis, MN). -

Evolutionary Divergence and Functions of the Human Interleukin (IL) Gene Family Chad Brocker,1 David Thompson,2 Akiko Matsumoto,1 Daniel W
UPDATE ON GENE COMPLETIONS AND ANNOTATIONS Evolutionary divergence and functions of the human interleukin (IL) gene family Chad Brocker,1 David Thompson,2 Akiko Matsumoto,1 Daniel W. Nebert3* and Vasilis Vasiliou1 1Molecular Toxicology and Environmental Health Sciences Program, Department of Pharmaceutical Sciences, University of Colorado Denver, Aurora, CO 80045, USA 2Department of Clinical Pharmacy, University of Colorado Denver, Aurora, CO 80045, USA 3Department of Environmental Health and Center for Environmental Genetics (CEG), University of Cincinnati Medical Center, Cincinnati, OH 45267–0056, USA *Correspondence to: Tel: þ1 513 821 4664; Fax: þ1 513 558 0925; E-mail: [email protected]; [email protected] Date received (in revised form): 22nd September 2010 Abstract Cytokines play a very important role in nearly all aspects of inflammation and immunity. The term ‘interleukin’ (IL) has been used to describe a group of cytokines with complex immunomodulatory functions — including cell proliferation, maturation, migration and adhesion. These cytokines also play an important role in immune cell differentiation and activation. Determining the exact function of a particular cytokine is complicated by the influence of the producing cell type, the responding cell type and the phase of the immune response. ILs can also have pro- and anti-inflammatory effects, further complicating their characterisation. These molecules are under constant pressure to evolve due to continual competition between the host’s immune system and infecting organisms; as such, ILs have undergone significant evolution. This has resulted in little amino acid conservation between orthologous proteins, which further complicates the gene family organisation. Within the literature there are a number of overlapping nomenclature and classification systems derived from biological function, receptor-binding properties and originating cell type. -

Open Chen-Thesis Finalv5.Pdf
The Pennsylvania State University The Graduate School Department of Neural and Behavioral Sciences EPIGENETIC ANALYSIS OF IMMUNE ASSOCIATED SIGNALING MOLECULES DURING MOUSE RETINA DEVELOPMENT A Thesis in Anatomy by Chen Yang © 2013 Chen Yang Submitted in Partial Fulfillment of the Requirements for the Degree of Master of Science May 2013 The thesis of Chen Yang was reviewed and approved* by the following: Samuel Shao-Min Zhang Assistant Professor of Neural and Behavioral Sciences Thesis Advisor Colin J. Barnstable Department Head of Neural and Behavioral Sciences Professor of Neural and Behavioral Sciences Patricia J. McLaughlin Professor of Neural and Behavioral Sciences Director of Graduate Program in Anatomy *Signatures are on file in the Graduate School. ii ABSTRACT The retina is an immune-privileged organ. Many autoimmune diseases, such as AMD, glaucoma, and diabetic retinopathy, are caused by excessive inflammatory responses targeting self-tissue. The physiological functions of extracellular and intracellular signaling molecules of immune responses have been well characterized. The epigenetic aspects of these molecules in the retina, however, have not been well elucidated. In this study, we examined the expression of selected immune-related genes, and their transcriptional accessibility via epigenetic mapping, cluster analysis, and RT-PCR. Among these genes, interleukin receptor related genes and intracellular signaling molecules exhibit higher transcriptional accessibility. Epigenetic mapping of the toll-like receptor (TLR) family revealed that 3 out of 13 TLRs exhibit H3K4me2 accumulation during retina development, suggesting that TLR2, TLR3, and TLR9 are the only TLR members expressed in the retina. Most of the NF-κB signaling molecules exhibited transcriptional accessibility, implying their essential roles in inflammatory regulation during retina maturation. -

Glycosylation of Immune Receptors in Cancer
cells Review Glycosylation of Immune Receptors in Cancer Ruoxuan Sun, Alyssa Min Jung Kim and Seung-Oe Lim * Department of Medicinal Chemistry and Molecular Pharmacology, Purdue Institute of Drug Discovery, Purdue Center for Cancer Research, Purdue University, West Lafayette, IN 47907, USA; [email protected] (R.S.); [email protected] (A.M.J.K.) * Correspondence: [email protected]; Tel.: +1-765-494-3531 Abstract: Evading host immune surveillance is one of the hallmarks of cancer. Immune checkpoint therapy, which aims to eliminate cancer progression by reprogramming the antitumor immune response, currently occupies a solid position in the rapidly expanding arsenal of cancer therapy. As most immune checkpoints are membrane glycoproteins, mounting attention is drawn to asking how protein glycosylation affects immune function. The answers to this fundamental question will stimulate the rational development of future cancer diagnostics and therapeutic strategies. Keywords: glycosylation; N-glycan; immune receptor; immune checkpoint therapy; cancer 1. Introduction The immune system can eliminate malignancy at an early stage by recognizing the antigenic peptide epitope presented by neoplastically transformed cells [1,2]. However, a subset of tumor cells may evolve to coexist with antitumor immunity by a process defined as immunoediting [3]. To combat the immune attack and maintain the balance with the host, cancer cells develop a series of approaches such as (1) downregulation of antigen Citation: Sun, R.; Kim, A.M.J.; Lim, presentation to make them invisible to immunity/the immune system, (2) production S.-O. Glycosylation of Immune of immunomodulatory cytokines to establish an immune suppressive environment, and Receptors in Cancer. Cells 2021, 10, (3) expression of surface immune checkpoint proteins such as programmed cell death 1 1100. -

Structure and Mechanism of Receptor Sharing by the IL-10R2 Common Chain
Structure Article Structure and Mechanism of Receptor Sharing by the IL-10R2 Common Chain Sung-il Yoon,1,4 Brandi C. Jones,1 Naomi J. Logsdon,1 Bethany D. Harris,2 Ashlesha Deshpande,1 Svetlana Radaeva,3 Brian A. Halloran,1 Bin Gao,3 and Mark R. Walter1,2,* 1Department of Microbiology 2Center for Biophysical Sciences and Engineering University of Alabama at Birmingham, Birmingham, AL 35294, USA 3Section on Liver Biology, Laboratory of Physiologic Studies, National Institute on Alcohol Abuse and Alcoholism, National Institutes of Health, Bethesda, MD 20892, USA 4Present address: Department of Molecular Biology, BCC-206, The Scripps Research Institute, 10550 North Torrey Pines Road, La Jolla, CA 92037, USA *Correspondence: [email protected] DOI 10.1016/j.str.2010.02.009 SUMMARY pard et al., 2003) heterodimers that are activated by IL-22, IL-26, and IL-28/ IL-29, respectively (see Figure S1 available IL-10R2 is a shared cell surface receptor required online). Thus, IL-10R2 functions as a common signaling chain for the activation of five class 2 cytokines (IL-10, in the class 2 cytokine family, as observed for gp130, IL-2gc IL-22, IL-26, IL-28, and IL-29) that play critical roles (gc), and GM-CSF bc chains in the class 1 cytokine family in host defense. To define the molecular mechanisms (Wang et al., 2009). that regulate its promiscuous binding, we have Cytokines that signal via IL-10R2 induce pleiotropic activities determined the crystal structure of the IL-10R2 ecto- that protect the host from overexuberant immune responses and activate innate immunity programs in epithelial cells. -

Proinflammatory Cytokine Activation Is Linked to Apoptotic Mediator, Soluble Fas Level in Patients with Chronic Heart Failure
Proinflammatory Cytokine Activation Is Linked to Apoptotic Mediator, Soluble Fas Level in Patients With Chronic Heart Failure Toru Kinugawa,1 MD, Masahiko Kato,2 MD, Kazuhiro Yamamoto,2 MD, Ichiro Hisatome,3 MD, and Ryuji Nohara,4 MD Summary The Fas/Fas Ligand system is a major apoptosis signaling pathway that is up-regulated in patients with chronic heart failure (CHF). Serum soluble Fas (sFas) levels increase in proportion to the CHF severity and may have prognostic value, therefore, sFas is a promising biomarker of heart failure. In this study, we attempted to identify the determinants of sFas levels in patients with CHF. Serum levels of tumor necrosis factor (TNF)-α and its soluble receptors (sTNF-R1 & sTNF-R2), interleukin (IL)-6, soluble IL-6 receptor (sIL-6R), glycoprotein (gp)130, and sFas were measured in 106 pa- tients with CHF and 39 controls. All subjects performed a symptom-limited cycle ergometer exercise test with expired gas analysis. CHF patients had higher levels of TNF-α, sTNF-R1, sTNF-R2, IL-6, and gp130. Serum levels of sFas (con- trols versus CHF; 2.60 ± 0.88 versus 3.38 ± 1.23 ng/mL, P = 0.0004) were higher in CHF. On univariate analysis, age (P = 0.0003), NYHA functional class (P = 0.0012), peak VO2 (P < 0.0001), plasma norepinephrine (P = 0.0013), log IL-6 (P < 0.0001), log TNF-α (P = 0.0002), log sTNF-R1 (P < 0.0001), and log TNF-R2 (P < 0.0001) were significantly re- lated to log sFas levels. Multivariate analysis showed that age and log IL-6 and log sTNF-R1 levels were independently associated with log sFas levels (overall R = 0.603, P < 0.0001). -

Signal-Transducing Receptor for IL-27 WSX-1 and Glycoprotein 130 Constitute A
WSX-1 and Glycoprotein 130 Constitute a Signal-Transducing Receptor for IL-27 Stefan Pflanz, Linda Hibbert, Jeanine Mattson, Rency Rosales, Elena Vaisberg, J. Fernando Bazan, Joseph H. This information is current as Phillips, Terrill K. McClanahan, Rene de Waal Malefyt and of October 1, 2021. Robert A. Kastelein J Immunol 2004; 172:2225-2231; ; doi: 10.4049/jimmunol.172.4.2225 http://www.jimmunol.org/content/172/4/2225 Downloaded from References This article cites 25 articles, 8 of which you can access for free at: http://www.jimmunol.org/content/172/4/2225.full#ref-list-1 http://www.jimmunol.org/ Why The JI? Submit online. • Rapid Reviews! 30 days* from submission to initial decision • No Triage! Every submission reviewed by practicing scientists • Fast Publication! 4 weeks from acceptance to publication by guest on October 1, 2021 *average Subscription Information about subscribing to The Journal of Immunology is online at: http://jimmunol.org/subscription Permissions Submit copyright permission requests at: http://www.aai.org/About/Publications/JI/copyright.html Email Alerts Receive free email-alerts when new articles cite this article. Sign up at: http://jimmunol.org/alerts The Journal of Immunology is published twice each month by The American Association of Immunologists, Inc., 1451 Rockville Pike, Suite 650, Rockville, MD 20852 Copyright © 2004 by The American Association of Immunologists All rights reserved. Print ISSN: 0022-1767 Online ISSN: 1550-6606. The Journal of Immunology WSX-1 and Glycoprotein 130 Constitute a Signal-Transducing Receptor for IL-27 Stefan Pflanz, Linda Hibbert, Jeanine Mattson, Rency Rosales, Elena Vaisberg, J. -
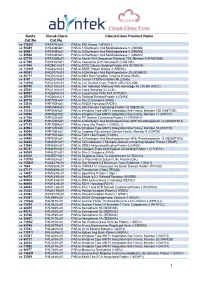
1 Santa Cat.No Cloud-Clone Cat.No. Cloud-Clone Product Name
Santa Cloud -Clone Cloud -Clone Product Name Cat.No Cat.No. sc -130253 PAS477Hu01 PAB to RIO Kinase 1 (RIOK1) sc -50485 PAS446Ra01 PAB to A Disintegrin And Metalloprotease 5 (ADAM5) sc -50487 PAS445Ra01 PAB to A Disintegrin And Metalloprotease 6 (ADAM6) sc -25588 PAS444Ra01 PAB to A Disintegrin And Metalloprotease 1 (ADAM1) sc -87719 PAR758Mu01 PAB to Family With Sequence Similarity 135, Member B (FAM135B) sc -67296 PAR493Hu01 PAB to Coenzyme Q10 Homolog B (COQ10B) sc -67048 PAQ981Hu01 PAB to S100 Calcium Binding Protein A15 (S100A15) sc -130269 PAQ342Hu01 PAB to SRSF Protein Kinase 3 (SRPK3) sc -98582 PAQ207Hu01 PAB to A Disintegrin And Metalloprotease 20 (ADAM20) sc -20711 PAQ164Hu01 PAB to BMX Non Receptor Tyrosine Kinase (BMX) sc -9147 PAQ127Hu01 PAB to Cluster Of Differentiation 8b (CD8b) sc -130184 PAQ124Hu01 PAB to Cell Division Cycle Protein 25B (CDC25B) sc -98790 PAQ118Hu01 PAB to Cell Adhesion Molecule With Homology To L1CAM (CHL1) sc -25361 PAQ116Hu01 PAB to Clock Homolog (CLOCK) sc -98937 PAQ089Hu01 PAB to Cytochrome P450 3A7 (CYP3A7) sc -25519 PAQ088Hu01 PAB to Dickkopf Related Protein 4 (DKK4) sc -28778 PAP797Hu01 PAB to Pim-2 Oncogene (PIM2) sc -33626 PAP750Hu01 PAB to RAD51 Homolog (RAD51) sc -8333 PAP694Mu01 PAB to SH2 Domain Containing Protein 1A (SH2D1A) sc -25524 PAP553Hu01 PAB to Wingless Type MMTV Integration Site Family, Member 10B (WNT10B) sc -50360 PAP552Hu01 PAB to Wingless Type MMTV Integration Site Family, Member 11 (WNT11) sc -87368 PAP332Hu01 PAB to PR Domain Containing Protein 14 (PRDM14) sc -25583 PAP226Hu01 -

Interleukin-6 Family Cytokines
Downloaded from http://cshperspectives.cshlp.org/ on October 5, 2021 - Published by Cold Spring Harbor Laboratory Press Interleukin-6 Family Cytokines Stefan Rose-John Institute of Biochemistry, Kiel University, Olshausenstrasse 40, Kiel, Germany Correspondence: [email protected] The interleukin (IL)-6 family cytokines is a group of cytokines consisting of IL-6, IL-11, ciliary neurotrophic factor (CNTF), leukemia inhibitory factor (LIF), oncostatin M (OSM), cardiotro- phin 1 (CT-1), cardiotrophin-like cytokine (CLC), and IL-27. They are grouped into one family because the receptorcomplex of each cytokine contains two (IL-6 and IL-11) or one molecule (all others cytokines) of the signaling receptor subunit gp130. IL-6 family cytokines have overlapping but also distinct biologic activities and are involved among others in the regula- tion of the hepatic acute phase reaction, in B-cell stimulation, in the regulation of the balance betweenregulatoryandeffectorTcells,inmetabolicregulation,andinmanyneuralfunctions. Blockade of IL-6 family cytokines has been shown to be beneficial in autoimmune diseases, but bacterial infections and metabolic side effects have been observed. Recent advances in cytokine blockade might help to minimize such side effects during therapeutic blockade. ytokines are small (15–20 kDa) and short- bopoietin, leukemia inhibitory factor (LIF), Clived proteins important in autocrine, para- and oncostatin M (OSM). Moreover, all inter- crine, and endocrine signaling. Cytokines coor- ferons and many colony-stimulating factors dinate the development and the activity of the (CSFs) belong to this class of cytokines, which immune system (Gandhi et al. 2016). Many cy- altogether contains far more than 60 members tokines belong to the four a-helical class of me- (Spangler et al.