Habitability Models for Planetary Sciences
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Where Less May Be More: How the Rare Biosphere Pulls Ecosystems Strings
The ISME Journal (2017) 11, 853–862 OPEN © 2017 International Society for Microbial Ecology All rights reserved 1751-7362/17 www.nature.com/ismej MINI REVIEW Where less may be more: how the rare biosphere pulls ecosystems strings Alexandre Jousset1, Christina Bienhold2,3, Antonis Chatzinotas4,5, Laure Gallien6,7, Angélique Gobet8, Viola Kurm9, Kirsten Küsel10,5, Matthias C Rillig11,12, Damian W Rivett13, Joana F Salles14, Marcel GA van der Heijden15,16,17, Noha H Youssef18, Xiaowei Zhang19, Zhong Wei20 and WH Gera Hol9 1Utrecht University, Department of Biology, Institute of Environmental Biology, Ecology and Biodiversity Group, Utrecht, The Netherlands; 2Alfred Wegener Institute Helmholtz Centre for Polar and Marine Research, Bremerhaven, Germany; 3Max Planck Institute for Marine Microbiology, Bremen, Germany; 4Helmholtz Centre for Environmental Research – UFZ, Leipzig, Germany; 5German Centre for Integrative Biodiversity Research (iDiv), Halle-Jena-Leipzig, Leipzig, Germany; 6Swiss Federal Institute for Forest, Snow and Landscape Research WSL, Birmensdorf, Switzerland; 7Center for Invasion Biology, Department of Botany & Zoology, Stellenbosch University, Matieland, South Africa; 8Sorbonne Universités, UPMC Université Paris 06, CNRS, UMR 8227, Integrative Biology of Marine Models, Station Biologique de Roscoff, F-29688, Roscoff Cedex, France; 9Netherlands Institute of Ecology, Department of Terrestrial Ecology, Wageningen, The Netherlands; 10Friedrich Schiller University Jena, Institute of Ecology, Jena, Germany; 11Freie Universtät Berlin, -

Molecular Biology for Computer Scientists
CHAPTER 1 Molecular Biology for Computer Scientists Lawrence Hunter “Computers are to biology what mathematics is to physics.” — Harold Morowitz One of the major challenges for computer scientists who wish to work in the domain of molecular biology is becoming conversant with the daunting intri- cacies of existing biological knowledge and its extensive technical vocabu- lary. Questions about the origin, function, and structure of living systems have been pursued by nearly all cultures throughout history, and the work of the last two generations has been particularly fruitful. The knowledge of liv- ing systems resulting from this research is far too detailed and complex for any one human to comprehend. An entire scientific career can be based in the study of a single biomolecule. Nevertheless, in the following pages, I attempt to provide enough background for a computer scientist to understand much of the biology discussed in this book. This chapter provides the briefest of overviews; I can only begin to convey the depth, variety, complexity and stunning beauty of the universe of living things. Much of what follows is not about molecular biology per se. In order to 2ARTIFICIAL INTELLIGENCE & MOLECULAR BIOLOGY explain what the molecules are doing, it is often necessary to use concepts involving, for example, cells, embryological development, or evolution. Bi- ology is frustratingly holistic. Events at one level can effect and be affected by events at very different levels of scale or time. Digesting a survey of the basic background material is a prerequisite for understanding the significance of the molecular biology that is described elsewhere in the book. -

Glossary Glossary
Glossary Glossary Albedo A measure of an object’s reflectivity. A pure white reflecting surface has an albedo of 1.0 (100%). A pitch-black, nonreflecting surface has an albedo of 0.0. The Moon is a fairly dark object with a combined albedo of 0.07 (reflecting 7% of the sunlight that falls upon it). The albedo range of the lunar maria is between 0.05 and 0.08. The brighter highlands have an albedo range from 0.09 to 0.15. Anorthosite Rocks rich in the mineral feldspar, making up much of the Moon’s bright highland regions. Aperture The diameter of a telescope’s objective lens or primary mirror. Apogee The point in the Moon’s orbit where it is furthest from the Earth. At apogee, the Moon can reach a maximum distance of 406,700 km from the Earth. Apollo The manned lunar program of the United States. Between July 1969 and December 1972, six Apollo missions landed on the Moon, allowing a total of 12 astronauts to explore its surface. Asteroid A minor planet. A large solid body of rock in orbit around the Sun. Banded crater A crater that displays dusky linear tracts on its inner walls and/or floor. 250 Basalt A dark, fine-grained volcanic rock, low in silicon, with a low viscosity. Basaltic material fills many of the Moon’s major basins, especially on the near side. Glossary Basin A very large circular impact structure (usually comprising multiple concentric rings) that usually displays some degree of flooding with lava. The largest and most conspicuous lava- flooded basins on the Moon are found on the near side, and most are filled to their outer edges with mare basalts. -

General Index
General Index Italicized page numbers indicate figures and tables. Color plates are in- cussed; full listings of authors’ works as cited in this volume may be dicated as “pl.” Color plates 1– 40 are in part 1 and plates 41–80 are found in the bibliographical index. in part 2. Authors are listed only when their ideas or works are dis- Aa, Pieter van der (1659–1733), 1338 of military cartography, 971 934 –39; Genoa, 864 –65; Low Coun- Aa River, pl.61, 1523 of nautical charts, 1069, 1424 tries, 1257 Aachen, 1241 printing’s impact on, 607–8 of Dutch hamlets, 1264 Abate, Agostino, 857–58, 864 –65 role of sources in, 66 –67 ecclesiastical subdivisions in, 1090, 1091 Abbeys. See also Cartularies; Monasteries of Russian maps, 1873 of forests, 50 maps: property, 50–51; water system, 43 standards of, 7 German maps in context of, 1224, 1225 plans: juridical uses of, pl.61, 1523–24, studies of, 505–8, 1258 n.53 map consciousness in, 636, 661–62 1525; Wildmore Fen (in psalter), 43– 44 of surveys, 505–8, 708, 1435–36 maps in: cadastral (See Cadastral maps); Abbreviations, 1897, 1899 of town models, 489 central Italy, 909–15; characteristics of, Abreu, Lisuarte de, 1019 Acequia Imperial de Aragón, 507 874 –75, 880 –82; coloring of, 1499, Abruzzi River, 547, 570 Acerra, 951 1588; East-Central Europe, 1806, 1808; Absolutism, 831, 833, 835–36 Ackerman, James S., 427 n.2 England, 50 –51, 1595, 1599, 1603, See also Sovereigns and monarchs Aconcio, Jacopo (d. 1566), 1611 1615, 1629, 1720; France, 1497–1500, Abstraction Acosta, José de (1539–1600), 1235 1501; humanism linked to, 909–10; in- in bird’s-eye views, 688 Acquaviva, Andrea Matteo (d. -

Bayesian Analysis of the Astrobiological Implications of Life's
Bayesian analysis of the astrobiological implications of life's early emergence on Earth David S. Spiegel ∗ y, Edwin L. Turner y z ∗Institute for Advanced Study, Princeton, NJ 08540,yDept. of Astrophysical Sciences, Princeton Univ., Princeton, NJ 08544, USA, and zInstitute for the Physics and Mathematics of the Universe, The Univ. of Tokyo, Kashiwa 227-8568, Japan Submitted to Proceedings of the National Academy of Sciences of the United States of America Life arose on Earth sometime in the first few hundred million years Any inferences about the probability of life arising (given after the young planet had cooled to the point that it could support the conditions present on the early Earth) must be informed water-based organisms on its surface. The early emergence of life by how long it took for the first living creatures to evolve. By on Earth has been taken as evidence that the probability of abiogen- definition, improbable events generally happen infrequently. esis is high, if starting from young-Earth-like conditions. We revisit It follows that the duration between events provides a metric this argument quantitatively in a Bayesian statistical framework. By (however imperfect) of the probability or rate of the events. constructing a simple model of the probability of abiogenesis, we calculate a Bayesian estimate of its posterior probability, given the The time-span between when Earth achieved pre-biotic condi- data that life emerged fairly early in Earth's history and that, billions tions suitable for abiogenesis plus generally habitable climatic of years later, curious creatures noted this fact and considered its conditions [5, 6, 7] and when life first arose, therefore, seems implications. -

Department of Pharmacology (GRAD) 1
Department of Pharmacology (GRAD) 1 faculty participate fully at all levels. The department has the highest DEPARTMENT OF level of NIH funding of all pharmacology departments and a great diversity of research areas is available to trainees. These areas PHARMACOLOGY (GRAD) include cell surface receptors, G proteins, protein kinases, and signal transduction mechanisms; neuropharmacology; nucleic acids, cancer, Contact Information and antimicrobial pharmacology; and experimental therapeutics. Cell and molecular approaches are particularly strong, but systems-level research Department of Pharmacology such as behavioral pharmacology and analysis of knock-in and knock-out Visit Program Website (http://www.med.unc.edu/pharm/) mice is also well-represented. Excellent physical facilities are available for all research areas. Henrik Dohlman, Chair Students completing the training program will have acquired basic The Department of Pharmacology offers a program of study that leads knowledge of pharmacology and related fields, in-depth knowledge in to the degree of doctor of philosophy in pharmacology. The curriculum is their dissertation research area, the ability to evaluate scientific literature, individualized in recognition of the diverse backgrounds and interests of mastery of a variety of laboratory procedures, skill in planning and students and the broad scope of the discipline of pharmacology. executing an important research project in pharmacology, and the ability The department offers a variety of research areas including to communicate results, analysis, and interpretation. These skills provide a sound basis for successful scientific careers in academia, government, 1. Receptors and signal transduction or industry. 2. Ion channels To apply to BBSP, students must use The Graduate School's online 3. -

The Search for Another Earth – Part II
GENERAL ARTICLE The Search for Another Earth – Part II Sujan Sengupta In the first part, we discussed the various methods for the detection of planets outside the solar system known as the exoplanets. In this part, we will describe various kinds of exoplanets. The habitable planets discovered so far and the present status of our search for a habitable planet similar to the Earth will also be discussed. Sujan Sengupta is an 1. Introduction astrophysicist at Indian Institute of Astrophysics, Bengaluru. He works on the The first confirmed exoplanet around a solar type of star, 51 Pe- detection, characterisation 1 gasi b was discovered in 1995 using the radial velocity method. and habitability of extra-solar Subsequently, a large number of exoplanets were discovered by planets and extra-solar this method, and a few were discovered using transit and gravi- moons. tational lensing methods. Ground-based telescopes were used for these discoveries and the search region was confined to about 300 light-years from the Earth. On December 27, 2006, the European Space Agency launched 1The movement of the star a space telescope called CoRoT (Convection, Rotation and plan- towards the observer due to etary Transits) and on March 6, 2009, NASA launched another the gravitational effect of the space telescope called Kepler2 to hunt for exoplanets. Conse- planet. See Sujan Sengupta, The Search for Another Earth, quently, the search extended to about 3000 light-years. Both Resonance, Vol.21, No.7, these telescopes used the transit method in order to detect exo- pp.641–652, 2016. planets. Although Kepler’s field of view was only 105 square de- grees along the Cygnus arm of the Milky Way Galaxy, it detected a whooping 2326 exoplanets out of a total 3493 discovered till 2Kepler Telescope has a pri- date. -

Amplification by PCR Artificially Reduces the Proportion of the Rare Biosphere in Microbial Communities
Amplification by PCR Artificially Reduces the Proportion of the Rare Biosphere in Microbial Communities Juan M. Gonzalez1*, Maria C. Portillo1, Pedro Belda-Ferre2, Alex Mira2 1 Institute of Natural Resources and Agrobiology, Spanish Council for Research, IRNAS-CSIC, Sevilla, Spain, 2 Genomics and Health Department, Center for Advanced Research in Public Health (CSISP), Valencia, Spain Abstract The microbial world has been shown to hold an unimaginable diversity. The use of rRNA genes and PCR amplification to assess microbial community structure and diversity present biases that need to be analyzed in order to understand the risks involved in those estimates. Herein, we show that PCR amplification of specific sequence targets within a community depends on the fractions that those sequences represent to the total DNA template. Using quantitative, real-time, multiplex PCR and specific Taqman probes, the amplification of 16S rRNA genes from four bacterial species within a laboratory community were monitored. Results indicate that the relative amplification efficiency for each bacterial species is a nonlinear function of the fraction that each of those taxa represent within a community or multispecies DNA template. Consequently, the low-proportion taxa in a community are under-represented during PCR-based surveys and a large number of sequences might need to be processed to detect some of the bacterial taxa within the ‘rare biosphere’. The structure of microbial communities from PCR-based surveys is clearly biased against low abundant taxa which are required to decipher the complete extent of microbial diversity in nature. Citation: Gonzalez JM, Portillo MC, Belda-Ferre P, Mira A (2012) Amplification by PCR Artificially Reduces the Proportion of the Rare Biosphere in Microbial Communities. -
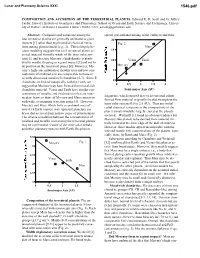
Composition and Accretion of the Terrestrial Planets
Lunar and Planetary Science XXXI 1546.pdf COMPOSITION AND ACCRETION OF THE TERRESTRIAL PLANETS. Edward R. D. Scott and G. Jeffrey Taylor, Hawai’i Institute of Geophysics and Planetology, School of Ocean and Earth Science and Technology, Univer- sity of Hawai’i at Manoa, Honolulu, Hawai’i 96822, USA; [email protected] Abstract: Compositional variations among the spread gravitational mixing of the embryos and their four terrestrial planets are generally attributed to giant 20 impacts [1] rather than to primordial chemical varia- Fig. 2 tions among planetesimals [e.g., 2]. This is largely be- Mars cause modeling suggests that each terrestrial planet ac- 15 creted material from the whole of the inner solar sys- tem [1], and because Mercury’s high density is attrib- 10 Venus Earth uted to mantle stripping in a giant impact [3] and not to its position as the innermost planet [4]. However, Mer- Mercury cury’s high concentration of metallic iron and low con- 5 centration of oxidized iron are comparable to those in recently discovered metal-rich chondrites [5-7]. Since E 0 chondrites are linked isotopically with the Earth, we 0 0.5 1 1.5 2 suggest that Mercury may have formed from metal-rich chondritic material. Venus and Earth have similar con- Semi-major Axis (AU) centrations of metallic and oxidized iron that are inter- fragments, which ensured that each terrestrial planet mediate between those of Mercury and Mars consistent formed from material originally located throughout the with wide, overlapping accretion zones [1]. However, inner solar system (0.5 to 2.5 AU). -

POSSIBLE STRUCTURE MODELS for the TRANSITING SUPER-EARTHS:KEPLER-10B and 11B
43rd Lunar and Planetary Science Conference (2012) 1290.pdf POSSIBLE STRUCTURE MODELS FOR THE TRANSITING SUPER-EARTHS:KEPLER-10b AND 11b. P. Futó1 1 Department of Physical Geography, University of West Hungary, Szombathely, Károlyi Gáspár tér, H- 9700, Hungary ([email protected]) Introduction:Up to january of 2012,10 super- The planet Kepler-11b has a large radius (1.97 R⊕) for Earths have been announced by Kepler-mission [1] its mass (4.3 M⊕),therefore this planet must have a that is designed to detect hundreds of transiting exo- spherical shell that is composed of low-density materi- planets.Kepler was launched on 6th March,2009 and the als.Considering the planet's average density,it must primary purpose of its scientific program is to search have a metallic core with different possible fractional for terrestrial-sized planets in the habitable zone of mass.Accordinghly,I have made a possible structure Solar-like stars.For the case of high number of discov- model for Kepler-11b in which the selected core mass eries we will be able to estimate the frequency of fraction is 32.59% (similarly to that of Earth) and the Earth-sized planets in our galaxy.Results of the Kepler- water ice layer has a relatively great fractional 's measurements show that the small-sized planets are volume.For case of the selected composition, the icy frequent in the spiral galaxies.A catalog of planetary surface sublimated to form a water vapor as the planet candidates,including objects with small-sized candidate moved inward the central star during its migration. -

The Subsurface Habitability of Small, Icy Exomoons J
A&A 636, A50 (2020) Astronomy https://doi.org/10.1051/0004-6361/201937035 & © ESO 2020 Astrophysics The subsurface habitability of small, icy exomoons J. N. K. Y. Tjoa1,?, M. Mueller1,2,3, and F. F. S. van der Tak1,2 1 Kapteyn Astronomical Institute, University of Groningen, Landleven 12, 9747 AD Groningen, The Netherlands e-mail: [email protected] 2 SRON Netherlands Institute for Space Research, Landleven 12, 9747 AD Groningen, The Netherlands 3 Leiden Observatory, Leiden University, Niels Bohrweg 2, 2300 RA Leiden, The Netherlands Received 1 November 2019 / Accepted 8 March 2020 ABSTRACT Context. Assuming our Solar System as typical, exomoons may outnumber exoplanets. If their habitability fraction is similar, they would thus constitute the largest portion of habitable real estate in the Universe. Icy moons in our Solar System, such as Europa and Enceladus, have already been shown to possess liquid water, a prerequisite for life on Earth. Aims. We intend to investigate under what thermal and orbital circumstances small, icy moons may sustain subsurface oceans and thus be “subsurface habitable”. We pay specific attention to tidal heating, which may keep a moon liquid far beyond the conservative habitable zone. Methods. We made use of a phenomenological approach to tidal heating. We computed the orbit averaged flux from both stellar and planetary (both thermal and reflected stellar) illumination. We then calculated subsurface temperatures depending on illumination and thermal conduction to the surface through the ice shell and an insulating layer of regolith. We adopted a conduction only model, ignoring volcanism and ice shell convection as an outlet for internal heat. -

2017 Journal Impact Factor (JCR)
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/317604703 2017 Journal Impact Factor (JCR) Technical Report · June 2017 CITATIONS READS 0 12,350 1 author: Pawel Domagala Pomeranian Medical University in Szczecin 34 PUBLICATIONS 326 CITATIONS SEE PROFILE All content following this page was uploaded by Pawel Domagala on 20 June 2017. The user has requested enhancement of the downloaded file. 1 , I , , 1 1 • • I , I • I : 1 t ( } THOMSON REUTERS - Journal Data Filtered By: Selected JCR Year: 2016 Selected Editions: SCIE,SSCI Selected Category Scheme: WoS Rank Full Journal Title Journal Impact Factor 1 CA-A CANCER JOURNAL FOR CLINICIANS 187.040 2 NEW ENGLAND JOURNAL OF MEDICINE 72.406 3 NATURE REVIEWS DRUG DISCOVERY 57.000 4 CHEMICAL REVIEWS 47.928 5 LANCET 47.831 6 NATURE REVIEWS MOLECULAR CELL BIOLOGY 46.602 7 JAMA-JOURNAL OF THE AMERICAN MEDICAL ASSOCIATION 44.405 8 NATURE BIOTECHNOLOGY 41.667 9 NATURE REVIEWS GENETICS 40.282 10 NATURE 40.137 11 NATURE REVIEWS IMMUNOLOGY 39.932 12 NATURE MATERIALS 39.737 13 Nature Nanotechnology 38.986 14 CHEMICAL SOCIETY REVIEWS 38.618 15 Nature Photonics 37.852 16 SCIENCE 37.205 17 NATURE REVIEWS CANCER 37.147 18 REVIEWS OF MODERN PHYSICS 36.917 19 LANCET ONCOLOGY 33.900 20 PROGRESS IN MATERIALS SCIENCE 31.140 21 Annual Review of Astronomy and Astrophysics 30.733 22 CELL 30.410 23 NATURE MEDICINE 29.886 24 Energy & Environmental Science 29.518 25 Living Reviews in Relativity 29.300 26 MATERIALS SCIENCE & ENGINEERING R-REPORTS 29.280 27 NATURE