The Lemming Cycle at Baker Lake, Northwest Territories, During 1959-62
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
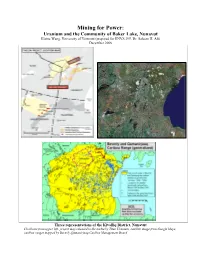
Mining for Power: Uranium Mining in Baker Lake Nunavut
Mining for Power: Uranium and the Community of Baker Lake, Nunavut Elaine Wang, University of Vermont (prepared for ENVS 295, Dr. Saleem H. Ali) December 2006 Three representations of the Kivalliq District, Nunavut Clockwise from upper left: project map released to the media by Titan Uranium; satellite image from Google Maps; caribou ranges mapped by Beverly-Qamanirjuaq Caribou Management Board “The Kiggavik Project near Baker Lake never went ahead. It is possible that, because of changing market conditions and the superior ore grades in neighboring Saskatchewan, 1989-90 had provided a window of opportunity for its development that was never to be repeated.” -Robert McPherson 2003, New Owners in the Own Land adapted from Cameco Corporation, 2006; not constant dollars Introduction In a plebiscite held on March 26, 1990, residents of the hamlet of Baker Lake (in what was then the District of Keewatin, Northwest Territories, now the District of Kivalliq, Nunavut, Canada), voted overwhelmingly against the development of a uranium mine by Urangesellschaft Canada Ltd. (UG) at a nearby site called Kiggavik, part of what prospectors know as the Thelon Basin. As a result, UG never explored its claims. In August of the same year, Bob Leonard, the president of the Keewatin Chamber of Commerce stated, “We are in an economic crisis. The economy in the Keewatin is in a mess. We are totally dependent on government spending and there’s no way that can continue.”1 The opening quote by Robert McPherson, a mining consultant in the Nunavut land claims negotiations, suggests that as recently as 2003, uranium mining near Baker Lake was, for many reasons, considered a non-option. -

Canadian Heritage Rivers System Management Plan for the Thelon River, N.W.T
CANADIAN HERITAGE RIVERS SYSTEM MANAGEMENT PLAN FOR THE THELON RIVER, N.W.T. Sector Tourism I 11-40.12 Plans/Strategies I I I - CANADIAN HERITAGE RIVERS SYSTEM MANAGEMENT PLAN FOR THE THELON RIVER, N.W.T. NWT EDT Can The 1990 — CANADIAN HERITAGE RIVERS SYSTEM MANAGEMENT PLAN FOR THE THELON RIVER, N. W.T. Submitted by the Municipality of Baker Lake; the Department of Economic Development and Tourism Government of the Northwest Territories; and the Department of Indian Affairs and Northern Development of Canada 1990 TABLE OF CONTENTS . 1.0 Introduction . 1.1 Thelon Heritage River Nomination . 1 1.2 Regional Setting and River Description . 1 1.3 Canadian Heritage Rivers System . 4 1.4 Purpose of the Management Plan . 4 l.4.1 General Considerations . 4 1.4.2 Objectives of the Thelon River Management Plan. 5 2.0 Background 2.1 History of the Nomination . 6 2.2 Public Support and Consultation . 6 2.3 Present Land Use . 8 2.3. lBaker Lake Inuit Land Use . 8 2.3.2 Land Tenure and Land Claims . 8 2.3.3 Mining and Other Development . 10 3.0 Heritage Values 3.1 Natural Heritage Values . 11 3.2 Human Heritage Values . 12 3.3 Recreational Values . 13 4.0 Planning and Management Program 4.1 Land Use Framework.. ~ . 15 4.1. 1 River Corridor . 15 4.1.2 Areas of Significance . 15 4.1.3 Potential Territorial Parks . 18 4.2 Heritage Management and Protection . 20 4.2.1 Human Heritage . 20 4.2.2 Natural Heritage . 21 4.2. -

A Roadmap to the Truth and Reconciliation Commission Call to Action #66
INDIGENOUS YOUTH VOICES A Roadmap to the Truth and Reconciliation Commission Call to Action #66 JUNE 2018 INDIGENOUS YOUTH VOICES 1 Kitinanaskimotin / Qujannamiik / Marcee / Miigwech: To all the Indigenous youth and organizations who took the time to share their ideas, experiences, and perspectives with us. To Assembly of Se7en Generations (A7G) who provided Indigenous Youth Voices Advisors administrative and capacity support. ANDRÉ BEAR GABRIELLE FAYANT To the Elders, mentors, friends and family who MAATALII ANERAQ OKALIK supported us on this journey. To the Indigenous Youth Voices team members who Citation contributed greatly to this Roadmap: Indigenous Youth Voices. (2018). A Roadmap to the Truth and Reconciliation Commission Call to Action #66. THEA BELANGER MARISSA MILLS Ottawa, Canada Anishinabe/Maliseet Southern Tuschonne/Michif Electronic ISBN Paper ISBN ERIN DONNELLY NATHALIA PIU OKALIK 9781550146585 9781550146592 Haida Inuk LINDSAY DUPRÉ CHARLOTTE QAMANIQ-MASON WEBSITE INSTAGRAM www.indigenousyouthvoices.com @indigenousyouthvoices Michif Inuk FACEBOOK TWITTER WILL LANDON CAITLIN TOLLEY www.fb.com/indigyouthvoices2 A ROADMAP TO TRC@indigyouthvoice #66 Anishinabe Algonquin ACKNOWLEDGMENT We would like to recognize and honour all of the generations of Indigenous youth who have come before us and especially those, who under extreme duress in the Residential School system, did what they could to preserve their language and culture. The voices of Indigenous youth captured throughout this Roadmap echo generations of Indigenous youth before who have spoken out similarly in hopes of a better future for our peoples. Change has not yet happened. We offer this Roadmap to once again, clearly and explicitly show that Indigenous youth are the experts of our own lives, capable of voicing our concerns, understanding our needs and leading change. -

Baker Lake HTO Presentation on Inuit Qaujimajatuqangit of Caribou Habitat
Baker Lake Hunters and Trappers Organization Presentation to Nunavut Wildlife Management Board Caribou Habitat Workshop November 4th, 2015 Iqaluit, Nunavut Basil Quinangnaq and Warren Bernauer Introduction • The Baker Lake Hunters and Trappers Organization (HTO) has requested mining and exploration activity be banned in caribou calving grounds and at important caribou water crossings. • This request is based on Inuit Qaujimajatuqangit – the traditional knowledge and teachings passed on from generations, and the wisdom and values of Baker Lake Elders • Baker Lake hunters and Elders have been fighting to protect these areas for decades • In September 2015, the Baker Lake HTO held a workshop with hunters and Elders, to discuss and share knowledge about these sensitive areas • In preparation for the workshop, Warren Bernauer conducted background research for the HTO. Warren wrote a report summarizing the documented knowledge of these areas, the importance of these areas to Inuit, and the long history of Baker Lake hunters fighting to protect these areas. Caribou Water Crossings • Caribou water crossings are some of the most important areas for caribou hunting for Baker Lake Inuit, both historically and today. • Caribou are very sensitive to disturbance at water crossings. • Inuit have many traditional rules for how water crossings should be treated. o Do not walk, hunt, skin animals, cache meat or camp on the side of the river where caribou enter the water. Even footprints will disturb caribou. o Camp upstream from water crossings; camps should not be visible from the crossing o Clean up all animal remains near a crossing. Even blood on the ground should be buried. -

Qikiqtani Region Arctic Ocean
OVERVIEW 2017 NUNAVUT MINERAL EXPLORATION, MINING & GEOSCIENCE QIKIQTANI REGION ARCTIC OCEAN OCÉAN ARCTIQUE LEGEND Commodity (Number of Properties) Base Metals, Active (2) Mine, Active (1) Diamonds, Active (2) Quttinirpaaq NP Sanikiluaq Mine, Inactive (2) Gold, Active (1) Areas with Surface and/or Subsurface Restrictions 10 CPMA Caribou Protection Measures Apply ISLANDS Belcher MBS Migratory Bird Sanctuary NP National Park Nares Strait Islands NWA National Wildlife Area - ÉLISABETH Nansen TP Territorial Park WP Wildlife Preserve WS Wildlife Sanctuary Sound ELLESMERE ELIZABETHREINE ISLAND Inuit Owned Lands (Fee simple title) Kane Surface Only LA Agassiz Basin Surface and Subsurface Ice Cap QUEEN Geological Mapping Programs Canada-Nunavut Geoscience Office ÎLES DE Kalaallit Nunaat Boundaries Peary Channel Müller GREENLAND/GROENLAND NLCA1 Nunavut Settlement Area Ice CapAXEL Nunavut Regions HEIBERG ÎLE (DENMARK/DANEMARK) NILCA 2 Nunavik Settlement Area ISLAND James Bay WP Provincial / Territorial D'ELLESMERE James Bay Transportation Routes Massey Sound Twin Islands WS Milne Inlet Tote Road / Proposed Rail Line Hassel Sound Prince of Wales Proposed Steensby Inlet Rail Line Prince Ellef Ringnes Icefield Gustaf Adolf Amund Meliadine Road Island Proposed Nunavut to Manitoba Road Sea Ringnes Eureka Sound Akimiski 1 Akimiski I. NLCA The Nunavut Land Claims Agreement Island Island MBS 2 NILCA The Nunavik Inuit Land Claims Agreement Norwegian Bay Baie James Boatswain Bay MBS ISLANDSHazen Strait Belcher Channel Byam Martin Channel Penny S Grise Fiord -

Arctic Science Day
Arctic Science Day An Introduction to Arctic Systems Science Research Conducted at the Centre for Earth Observation Science (U of MB) Produced by: Michelle Watts Schools on Board Program Coordinator Arctic Geography – a brief introduction The Arctic Region is the region around the North Pole, usually understood as the area within the Arctic Circle. It includes parts of Russia, Scandinavia, Greenland, Canada, Alaska and the Arctic Ocean. The Arctic is a single, highly integrated system comprised of a deep, ice covered and nearly isolated ocean surrounded by the land masses of Eurasia and North America. It is made up of a range of land-and seascapes from mountains and glaciers to flat plains, from coastal shallows to deep ocean basins, from polar deserts to sodden wetlands, from large rivers to isolated ponds. Sea ice, permafrost, glaciers, ice sheets, and river and lake ice are all characteristic parts of the Arctic’s physical geography (see circumpolar map) Inuit Regions of Canada (www.itk.ca) Inuit Regions of Canada – See Map Inuit Nuanagat There are four Inuit regions in Canada, collectively known as Inuit Nunangat. The term “Inuit Nunangat” is a Canadian Inuit term that includes land, water, and ice. Inuit consider the land, water, and ice, of our homeland to be integral to our culture and our way of life. Inuvialuit (Northwest Territories) The Inuvialuit region comprises the northwestern part of the Northwest Territories. In 1984, the Inuvialuit, federal and territorial governments settled a comprehensive land claims agreement, giving Inuvialuit surface and subsurface (mining) rights to most of the region. The Agreement ensures environmental protection, harvesting rights and Inuvialuit participation and support in many economic development initiatives. -

Community Experiences of Mining in Baker Lake, Nunavut
Community Experiences of Mining in Baker Lake, Nunavut by Kelsey Peterson A Thesis presented to The University of Guelph In partial fulfillment of requirements for the degree of Master of Arts in Geography Guelph, Ontario, Canada ©Kelsey Peterson, April, 2012 ABSTRACT COMMUNITY EXPERIENCES OF MINING IN BAKER LAKE, NUNAVUT Kelsey Peterson Advisor: University of Guelph, 2012 Professor Ben Bradshaw With recent increases in mineral prices, the Canadian Arctic has experienced a dramatic upswing in mining development. With the development of the Meadowbank gold mine, the nearby Hamlet of Baker Lake, Nunavut is experiencing these changes firsthand. In response to an invitation from the Hamlet of Baker Lake, this research document residents’ experiences with the Meadowbank mine. These experiences are not felt homogeneously across the community; indeed, residents’ experiences with mining have been mixed. Beyond this core finding, the research suggests four further notable insights: employment has provided the opportunity for people to elevate themselves out of welfare/social assistance; education has become more common, but some students are leaving high school to pursue mine work; and local businesses are benefiting from mining contracts, but this is limited to those companies pre-existing the mine. Finally, varied individual experiences are in part generated by an individual’s context including personal context and choices. ii ACKNOWLEDGEMENTS This thesis would not have been possible without the support and assistance of several key people. My advisor, Ben Bradshaw, has been instrumental in all stages of pre- research, research and writing. From accepting me to the program and connecting me to Baker Lake to funding my research and reading innumerable drafts of the written thesis, Professor Bradshaw has been critical to this research project. -

Inuit Ethnobotany and Ethnoecology in Nunavik and Nunatsiavut, Northeastern Canada
Université de Montréal Inuit ethnobotany and ethnoecology in Nunavik and Nunatsiavut, northeastern Canada par Courtenay Clark Département de sciences biologiques Faculté des arts et des sciences Mémoire présenté à la Faculté des études supérieures en vue de l’obtention du grade de maîtrise en sciences biologiques Décembre, 2012 © Courtenay Clark, 2012 a Université de Montréal Faculté des études supérieures Ce mémoire intitulé Inuit ethnobotany and ethnoecology in Nunavik and Nunatsiavut, northeastern Canada Présenté par : Courtenay Clark a été évalué par un jury composé des personnes suivantes : Stéphanie Pellerin Président-rapporteur Alain Cuerrier Directeur de recherche Thora Martina Herrmann Membre du jury b Résumé Les habitats uniques de l'écotone forêt boréale-subarctique dans le nord du Canada subissent les contrecoups du changement climatique. Combinés aux effets de la mondialisation, les changements environnementaux touchent les Inuits de cette région et imposent des contraintes importantes sur leur mode de vie traditionnel, ce qui a des répercussions sur leur langue et les savoirs qui l'accompagnent. Cette étude compare deux aspects de l’ethnobiologie inuite : a) les noms et les utilisations des plantes par les Inuits de Nain, Nunatsiavut, suivis par une comparaison des utilisations avec la communauté inuite de Kangiqsualujjuaq, Nunavik, et b) une analyse des types de lieux ou d’habitats que les Inuits reconnaissent et nomment. Des interviews semi-dirigés ont été menés à Nain, Nunatsiavut et à Kangiqsualujjuaq, au Nunavik. Les plantes mentionnées sont utilisées comme aliment, thé, médecine, combustible, construction, nettoyage, et autres utilisations. Les deux communautés ont utilisé un nombre égal de plantes, avec des proportions équivalentes de taxons vasculaires/invasculaires, de formes de croissance (habitus), et d’espèces par catégorie d'utilisation. -

Baker Lake 2009 Baker Lake, Or Qamani’Tuuq, Is Situated Inland, at the Huge Widening at the Mouth of the Thelon River, Close to the Geographic Centre of Canada
Baker Lake 2009 Baker Lake, or Qamani’tuuq, is situated inland, at the huge widening at the mouth of the Thelon River, close to the geographic centre of Canada. Baker Lake is well known for its arts and craft community. Distinguished artists and studios promoting arts and crafts, such as the Inuit Heritage Centre, the Jessie Oonark Centre, and well-established independent art galleries are assets to the community’s arts and craft industry. The natural resource industry could also have a significant impact on the economic development of Baker Lake. Further development of the Meawdowbank gold deposit, now at the initial review stages with their environmental impact statement, has already created numerous local hires. If mine proceeds into operation, more jobs will be available at the mine site, approximately 70kms from Baker Lake. From administration to cooking to heavy equipment operation, community council is now proactively looking into training programs to prepare its residents for the assortment of jobs that may be available. For more information about Baker Lake, visit www.bakerlake.org. Getting There: Calm Air operates daily flights from Winnipeg. Please check with the airline for schedule changes. (For larger map use zoom tool) Community Services and Information Population 1,728 Region Kivalliq Time Zone Central Postal Code X0C 0A0 Population based on 2006 Stats Canada Report Area Code is 867, unless as indicated. RCMP General Inquiries 793-0123 Local Communications Emergency Only 793-1111 Arctic Net (broadband) 793-4044 Health -

Satellite Microwave Assessment of Northern Hemisphere Lake Ice Phenology from 2002 to 2015“ by Jinyang Du, John S
Reply to RC2 for the manuscript “Satellite Microwave Assessment of Northern Hemisphere Lake Ice Phenology from 2002 to 2015“ by Jinyang Du, John S. Kimball, Claude Duguay, Youngwook Kim, and Jennifer D. Watts Dear Anonymous Referee #2, thank you for your helpful comments on our manuscript. Please find below our responses to all the comments (in bold and italic). The changes on the manuscript were highlighted in blue. This manuscript presents a new methodology to apply the use of passive microwave imagery (AMSR-E/2) to create an automated daily ice phenology product utilized for monitoring freshwater lake ice. The daily temporal ability, unhindered by cloud and polar darkness allows for a global product at a resolution and accuracy that can of benefit climate modelling and global change studies. This manuscript is well written and a valuable contribution to the freshwater ice research field. The inclusion of cited literature is light in the introduction section and could use some strengthening; however overall, the manuscript is clear, concise and suitable for publication with only minor revision to the text and figures. No particular scientific issues arise from this review. The validation of the new passive microwave based product is limited by the lack of existing observation data from lakes, however including the comparison to the CIS dataset and the IMS product strengthens the validation sufficiently. Reply: Thank you for the comments. The following is a list of some minor typographical suggestions and some points to clarify the figures: Line 11 : Previous studies have documented significantly earlier ice break-up between 1951 and 2000 for lakes in Canada (Duguay et al., 2006) and decreasing lake ice cover of Lake Ladoga in Europe during the last few decades (Karetnikov and Naumenko, 2008). -

First Canadians, Canadians First: National Strategy on Inuit
First Canadians, Canadians First NATIONAL STRATEGY ON INUIT EDUCATION 2011 FIRST CANADIANS, CANADIANS FIRST “Do Inuit see themselves as Inuit first or as Canadians first? I have always thought those two sentiments were one and the same. After all, during our many meetings with Inuit from countries such as Denmark, the United States or Russia, we have always been Canadian Inuit.” — Jose Kusugak “ᐃᓄᐃᑦ ᓇᖕᒥᓂᖅ ᐃᓱᒪᒋᕙᒃᐸᑦ ᐃᓅᓂᖏᑦ ᓯᕗᓪᓕᐅᑎᓪᓗᒍ ᐅᑉᐸᓘᑉᐸ ᑲᓇᑕᒥᐅᑕᐅᓂᖏᑦ ᓯᕗᓪᓕᐅᑎᓪᓗᒍ? ᑕᒪᒃᑯᐊᒃ ᒪᕐᕉᒃ ᐃᓱᒫᒃ ᐊᑕᐅᓯᐅᓱᒌᓐᓇᖅᓯᒪᔮᒃᑲᒃ ᐊᔾᔨᒌᖑᓪᓗᑎᒡᓗ. ᓲᖃᐃᒻᒪ, ᐅᓄᖅᑐᓂᒃ ᑲᑎᒪᓂᖃᖅᓯᒪᓕᖅᑎᓪᓗᑕ ᐃᓄᖕᓂᒃ ᓄᓇᓕᕐᔪᐊᕐᓂᙶᖅᑐᓂᒃ ᐃᒻᒪᖄ ᑕᓐᒫᒃᒥᑦ, ᐊᒥᐊᓕᑲᒃᑯᓐᓂᑦ ᐅᑉᐸᓘᓐᓃᑦ ᕋᓴᒥᑦ, ᑲᓇᑕᒥᐅᑕᐅᓪᓗᑕ ᐃᓅᓂᕋᐃᓐᓇᖅᓯᒪᔪᒍᑦ.” — ᔫᓯ ᐊᒪᐅᔭᖅ ᑯᓱᒐᖅ “Inuit inminik takuhimayut Inuuplutik hivulimik ovaluniit Kanatamiut hivulimik? Ihumainaktunga tahapkoa ihumaiyait atauhiuyut ovalo aatjikiikhutik. Ilaa, amigaitunik katimakataligaagapta Inuit aalanit nunait, ilangit; Denmark, Amialikat ovaluniit Rusiat, ilitagiinaktugut Kanatamiut Inuuyugut.” — Jose Kusugak “Inuit takunnausiKavât Inullutik sivullipâmi ubvalu Canada- miungullutik sivullimi? IsumaKainnalaukKunga maggok atausiulin- ganeginnik. Unuttuvatsuani katimaKatigennitinni Inunnut nunalinnit sollu Denmark, United States ubvalu Russian, Canada-miungulluta Inovugut.” — Jose Kusugak « Les Inuits se considèrent-ils comme Inuits ou Canadiens en premier lieu? J’ai toujours pensé que ces deux sentiments se confondent en un seul. Après tout, lors de nos nombreuses réunions avec les Inuits d’autres pays comme le Danemark, les États-Unis ou la Russie, nous avons toujours été considérés comme les Inuits canadiens. » — Jose Kusugak 1 FIRST CANADIANS, CANADIANS FIRST Jose Kusugak (1950-2011) was a leader, storyteller and a passionate advocate for Inuit rights. The National Committee on Inuit Education dedicates this National Strategy on Inuit Education to him for his contributions to Inuit education and the Inuit language. Through his insights, humour and understanding of the human condition, he gave us the gift of imagining the possible. -

Canada Archives Canada Published Heritage Direction Du Branch Patrimoine De I'edition
Uqaujjuusiat - Gifts of Words of Advice: Schooling, Education and Leadership in Baffin Island A Thesis Submitted to the Faculty of Education In Partial Fulfilment of the Requirements for the Degree of Master of Education University of Prince Edward Island We accept this thesis as conforming to the required standards Dr. Fiona Walton Dr. Alexander McAuley Naullaq Arnaquq Iqaluit, NU April, 2008 © 2008 Arnaquq Library and Bibliotheque et 1*1 Archives Canada Archives Canada Published Heritage Direction du Branch Patrimoine de I'edition 395 Wellington Street 395, rue Wellington Ottawa ON K1A0N4 Ottawa ON K1A0N4 Canada Canada Your file Votre reference ISBN: 978-0-494-49838-5 Our file Notre reference ISBN: 978-0-494-49838-5 NOTICE: AVIS: The author has granted a non L'auteur a accorde une licence non exclusive exclusive license allowing Library permettant a la Bibliotheque et Archives and Archives Canada to reproduce, Canada de reproduire, publier, archiver, publish, archive, preserve, conserve, sauvegarder, conserver, transmettre au public communicate to the public by par telecommunication ou par Plntemet, prefer, telecommunication or on the Internet, distribuer et vendre des theses partout dans loan, distribute and sell theses le monde, a des fins commerciales ou autres, worldwide, for commercial or non sur support microforme, papier, electronique commercial purposes, in microform, et/ou autres formats. paper, electronic and/or any other formats. The author retains copyright L'auteur conserve la propriete du droit d'auteur ownership and moral rights in et des droits moraux qui protege cette these. this thesis. Neither the thesis Ni la these ni des extraits substantiels de nor substantial extracts from it celle-ci ne doivent etre imprimes ou autrement may be printed or otherwise reproduits sans son autorisation.