GEOMORPHIC CONTROLS, RIFFLE SUBSTRATE QUALITY, and SPAWNING SITE SELECTION in TWO SEMI-ALLUVIAL SALMON RIVERS in the GASPE´ PENINSULA, Canaday
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Red River: the Northern Border of Texas!
“From Palo Duro Canyon outside Amarillo Texas The prairie dog town fork of the Red River flows Headed cross the plains along the coast of Oklahoma To the Mississippi River and the Gulf of Mexico” Red River by Guy Clark Red River: The northern Border of Texas! By Wm Davey Edwards, PhD Texas & Oklahoma Professional Land Surveyor Texas Licensed State Land Surveyor US Federal Land Surveyor Edwards Surveying, LLC Decatur, Texas Red River Boundary between Texas and Oklahoma South bank of the Red River in July 2015 the day after a rain. Objectives • Natural Boundaries • Accretion, Erosion, and Avulsion • BLM Claims • Conclusion Natural Boundaries for International Borders Adams – Onís Treaty of 1819 • Sabine, Red, and Arkansas Rivers • begin on the Gulf of Mexico, at the mouth of the River Sabine in the Sea, continuing North, along the Western Bank of that River, to the 32d degree of Latitude; thence by a Line due North to the degree of Latitude, where it strikes the Rio Roxo of Natchitoches, or Red-River, then following the course of the Rio-Roxo Westward to the degree of Longitude, 100 West from London and 23 from Washington, then crossing the said Red-River, and running thence by a Line due North to the River Arkansas, thence, following the Course of the Southern bank of the Arkansas to its source in Latitude, 42. North and thence by that parallel of Latitude to the South-Sea Caroline C. Stafford v. Adam C. King • SUPREME COURT OF TEXAS, 30 Tex. 257, April, 1867 Natural objects are mountains, lakes, rivers, creeks, rocks, and the like; artificial objects are marked trees, stakes, mounds, etc., constructed by others or the surveyor, and called for in the field- notes, and they should be inserted in the patent. -

Lesson 4: Sediment Deposition and River Structures
LESSON 4: SEDIMENT DEPOSITION AND RIVER STRUCTURES ESSENTIAL QUESTION: What combination of factors both natural and manmade is necessary for healthy river restoration and how does this enhance the sustainability of natural and human communities? GUIDING QUESTION: As rivers age and slow they deposit sediment and form sediment structures, how are sediments and sediment structures important to the river ecosystem? OVERVIEW: The focus of this lesson is the deposition and erosional effects of slow-moving water in low gradient areas. These “mature rivers” with decreasing gradient result in the settling and deposition of sediments and the formation sediment structures. The river’s fast-flowing zone, the thalweg, causes erosion of the river banks forming cliffs called cut-banks. On slower inside turns, sediment is deposited as point-bars. Where the gradient is particularly level, the river will branch into many separate channels that weave in and out, leaving gravel bar islands. Where two meanders meet, the river will straighten, leaving oxbow lakes in the former meander bends. TIME: One class period MATERIALS: . Lesson 4- Sediment Deposition and River Structures.pptx . Lesson 4a- Sediment Deposition and River Structures.pdf . StreamTable.pptx . StreamTable.pdf . Mass Wasting and Flash Floods.pptx . Mass Wasting and Flash Floods.pdf . Stream Table . Sand . Reflection Journal Pages (printable handout) . Vocabulary Notes (printable handout) PROCEDURE: 1. Review Essential Question and introduce Guiding Question. 2. Hand out first Reflection Journal page and have students take a minute to consider and respond to the questions then discuss responses and questions generated. 3. Handout and go over the Vocabulary Notes. Students will define the vocabulary words as they watch the PowerPoint Lesson. -

Topic 12 – Shaping the Earth Vocabulary
Topic 12 – Shaping the Earth Vocabulary Abrasion – physical action of scraping, rubbing, grinding or wearing away rock material Chemical weathering – the process of using natural chemical reactions to break down rock Cut bank – Outside bend in a stream where the water velocity is fastest and erosion is the greatest. Delta – the region of a stream mouth where sediment is deposited as it flows into a large body of water in graded beds of largest to smallest particles Deposition – the process where sediment is dropped out of the stream current and builds into layers Drumlin – a glacial feature that is created as a glacier flows. The blunt end shows the direction of flow Erosion – the removal of sediment and weather material from rock Glacier – a mass of ice that flows due to gravity Kettle lake – a small round lake that is formed when a large chunk of glacial ice creates a depression in the Earth’s surface and melts Landscape – regions on Earth’s surface with similar surface feature such as mountains, plains and plateaus Mass movement – the movement of large quantities of earth materials due to gravity, includes landslides, mudslides, rock slides and soil creep Meander – the curve or bend of a stream channel Moraine – a large ridge, pile or sheet of unsorted sediment deposited by a glacier Outwash plain – a glacial feature of sorted layered sediment as the result of the glacier melting Physical weathering – the mechanical breakdown of rock material at the earth’s surface into smaller pieces Point bar – Opposite the cut bank in a stream where the velocity is slowest and deposition is the greatest. -
Montana State Parks Guide Reservations for Camping and Other Accommodations: Toll Free: 1-855-922-6768 Stateparks.Mt.Gov
For more information about Montana State Parks: 406-444-3750 TDD: 406-444-1200 website: stateparks.mt.gov P.O. Box 200701 • Helena, MT 59620-0701 Montana State Parks Guide Reservations for camping and other accommodations: Toll Free: 1-855-922-6768 stateparks.mt.gov For general travel information: 1-800-VISIT-MT (1-800-847-4868) www.visitmt.com Join us on Twitter, Facebook & Instagram If you need emergency assistance, call 911. To report vandalism or other park violations, call 1-800-TIP-MONT (1-800-847-6668). Your call can be anonymous. You may be eligible for a reward. Montana Fish, Wildlife & Parks strives to ensure its programs, sites and facilities are accessible to all people, including those with disabilities. To learn more, or to request accommodations, call 406-444-3750. Cover photo by Jason Savage Photography Lewis and Clark portrait reproductions courtesy of Independence National Historic Park Library, Philadelphia, PA. This document was produced by Montana Fish Wildlife & Parks and was printed at state expense. Information on the cost of this publication can be obtained by contacting Montana State Parks. Printed on Recycled Paper © 2018 Montana State Parks MSP Brochure Cover 15.indd 1 7/13/2018 9:40:43 AM 1 Whitefish Lake 6 15 24 33 First Peoples Buffalo Jump* 42 Tongue River Reservoir Logan BeTableaverta ilof Hill Contents Lewis & Clark Caverns Les Mason* 7 16 25 34 43 Thompson Falls Fort3-9 Owen*Historical Sites 28. VisitorMadison Centers, Buff Camping,alo Ju mp* Giant Springs* Medicine Rocks Whitefish Lake 8 Fish Creek 17 Granite11-15 *Nature Parks 26DisabledMissouri Access Headw ibility aters 35 Ackley Lake 44 Pirogue Island* WATERTON-GLACIER INTERNATIONAL 2 Lone Pine* PEACE PARK9 Council Grove* 18 Lost Creek 27 Elkhorn* 36 Greycliff Prairie Dog Town* 45 Makoshika Y a WHITEFISH < 16-23 Water-based Recreation 29. -

The Spatial Distribution of Bed Sediment on Fluvial System: a Mini Review of the Aceh Meandering River
Aceh Int. J. Sci. Technol., 5(2): 82-87 August 2016 doi: 10.13170/aijst.5.2.4932 Aceh International Journal of Science and Technology ISSN: 2088-9860 Journal homepage: http://jurnal.unsyiah.ac.id/aijst The Spatial Distribution of Bed Sediment on Fluvial System: A Mini Review of the Aceh Meandering River Muhammad Irham Faculty of Marine and Fisheries Science, University of Syiah Kuala, Banda Aceh 23111, Indonesia. Corresponding author, email: [email protected] Received : 2 August 2016 Accepted : 28 August 2016 Online : 31 August2016 Abstract - Dynamic interactions of hydrological and geomorphological processes in the fluvial system result in accumulated deposit on the bed because the capacity to carry sediment has been exceeded. The bed load of the Aceh fluvial system is primarily generated by mechanical weathering resulting in boulders, pebbles, and sand, which roll or bounce along the river bed forming temporary deposits as bars on the insides of meander bends, as a result of a loss of transport energy in the system. This dynamic controls the style and range of deposits in the Aceh River. This study focuses on the spatial distribution of bed-load transport of the Aceh River. Understanding the spatial distribution of deposits facilitates the reconstruction of the changes in controlling factors during accumulation of deposits. One of the methods can be done by sieve analysis of sediment, where the method illuminates the distribution of sediment changes associate with channel morphology under different flow regimes. Hence, the purpose of this mini review is to investigate how the sediment along the river meander spatially dispersed. -
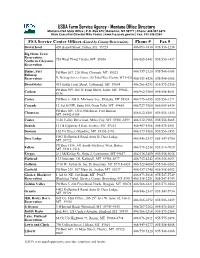
PDF of Montana
USDA Farm Service Agency - Montana Office Directory Montana FSA State Office | P.O. Box 670 | Bozeman, MT 59771 | Phone: 406.587.6872 State Executive Director Mike Foster | www.fsa.usda.gov/mt | Fax: 855.546.0264 FSA Service Center Offices (Listed by County/Reservation) Phone # Fax # Beaverhead 420 Barrett Street, Dillon, MT 59725 406/683-3830 855/556-1258 Big Horn, Crow Reservation, 724 West Third, Hardin, MT 59034 406/665-3442 855/556-1457 Northern Cheyenne Reservation Blaine, Fort PO Box 307, 228 Ohio, Chinook, MT 59523 406/357-2320 855/546-0388 Belknap Reservation Ft. Belknap Service Center, 158 Tribal Way, Harlem, MT 59526 406/353-8526 855/546-0388 Broadwater 415 South Front Street, Townsend, MT 59644 406/266-4253 855/575-2506 PO Box 509, 606 W Front Street, Joliet, MT 59041- Carbon 406/962-3300 855/558-5641 0136 Carter PO Box 5, 308 S. Mormon Ave., Ekalaka, MT 59324 406/775-6355 855/556-1271 Cascade 12 3rd St NW, Suite 300, Great Falls, MT 59404 406/727-7580 866/609-8434 PO Box 309, 1210 25th Street, Fort Benton, Chouteau 406/622-5401 855/556-1450 MT 59442-0309 Custer 3120 Valley Drive East, Miles City, MT 59301-5599 406/232-7905 855/558-5665 Daniels 131 B Highway 5 East, Scobey, MT 59263 406/487-5366 855/575-2501 Dawson 102 Fir Street, Glendive, MT 59330-3196 406/377-5566 855/556-1455 1002 Hollenback Road, Suite B, Deer Lodge, Deer Lodge 406/846-2337 855-547-5750 MT 59722 PO Box 1516, 141 South 4th Street West, Baker, Fallon 406/778-2238 855-510-7029 MT 59313-1516 Fergus 211 McKinley St., Suite 2, Lewistown, MT 59457 406/538-3489 855/558-5654 Flathead 133 Interstate LN, Kalispell, MT 59901-2877 406/752-4242 855/558-5653 Gallatin 3710 W. -

Glacier County Pre-Disaster Mitigation Plan February 2017
Glacier County Pre-Disaster Mitigation Plan February 2017 Encompassing the Jurisdictions of: Glacier County City of Cut Bank Prepared By: Fulton Wold, CBCP Tony Gertz, AEM Executive Officer Mitigation Planner (615) 469-5558 (615) 469-5558 [email protected] [email protected] Glacier County Pre-Disaster Mitigation Plan Page 2 Executive Summary The Glacier County Pre-Disaster Mitigation Plan is being developed to update and revise hazard mitigation activities for Glacier County. The Glacier County Mitigation Planning Committee will evaluate mitigation measures to be undertaken, and outline a strategy for implementation of mitigation projects. This plan covers two municipalities in Glacier County, including the governments of: Glacier County and the City of Cut Bank. Formal adoption and implementation of a pre-disaster mitigation plan may present many benefits to Glacier County and Cut Bank. By identifying problems and possible solutions in advance of a disaster, Glacier County and Cut Bank will be in a better position to obtain pre and post-disaster funding. This document aims to produce the following strategic outcomes: 1) Reduce loss of life and decrease property losses to Glacier County and its jurisdictions due to natural disasters; and 2) Provide the framework and coordination to encourage government, and both public and private organizations at all levels, to undertake mitigation in order to minimize potential disasters and to employ mitigation strategies in the recovery following disasters. Specifically, these strategic -

City of Cut Bank Public Water Supply PWSID # MT0000193
City of Cut Bank Public Water Supply PWSID # MT0000193 Date of Report: March 20, 2003 SOURCE WATER DELINEATION AND ASSESSMENT REPORT Operator/Manager: Keith Nelson 113 East Main Street Cut Bank, MT 59427 (406) 873-2362 City of Cut Bank SWDAR Table of Contents INTRODUCTION .....................................................................................................................................................................1 PURPOSE...............................................................................................................................................................................1 LIMITATIONS.......................................................................................................................................................................1 BACKGROUND........................................................................................................................................................................2 THE COMMUNITY...............................................................................................................................................................2 CLIMATE...............................................................................................................................................................................2 GEOGRAPHIC SETTING .....................................................................................................................................................3 GEOLOGY .............................................................................................................................................................................3 -

Glacier Area Emergency Response Action Plan
GLACIER AREA EMERGENCY RESPONSE ACTION PLAN PHMSA Sequence Number 2987 EPA FRP ID Number (Pending) Owner/Operator: Phillips 66 2331 City West Blvd. Houston, Texas 77042 24-Hour Number: (800) 231-2551 or (877) 267-2290 Page A7-1 Confidentiality Notice: This document is for the sole use of the intended recipient(s) and contains information that is considered to be proprietary to Phillips 66. Any unauthorized review, use, disclosure or distribution is strictly prohibited. Glacier Area Appendix 7: Response Zone Appendix ERAP Appendix 7: ERAP Table of Contents 7.1 Area Information ........................................................................................................1 7.1.1 Glacier Crude North .......................................................................................1 7.1.2 Cut Bank Station ............................................................................................ 4 7.1.3 Tank Table .....................................................................................................5 7.2 Communication Equipment ........................................................................................6 7.3 Notification Sequence ................................................................................................7 7.4 Emergency Notification Contact List ........................................................................ 10 7.5 Emergency Response Equipment, Testing & Deployment ....................................... 13 7.6 Evacuation Plan ...................................................................................................... -

Geomorphic Evaluation of Little Topashaw Creek: June 2000
Geomorphic Evaluation of Little Topashaw Creek: June 2000 N. Wallerstein Edited by F. Douglas Shields, Jr. 1 Introduction This brief report outlines observations made on the geomorphology of Little Topashaw Creek in June 2000. The reason for this analysis is to give a baseline survey of the reach prior to the installation of large woody debris structures which will be placed in the outside of the apex of several bends to protect the banks from fluvial erosion. The reach in question is located immediately north of the bridge carrying a county road along the Chickasaw-Webster county line and is approximately 2000 meters in length. Thirty-nine cross sections were surveyed in the reach between 1999 and January 2000. Cross sections 1-27, 29, 31,33, 35,37 & 39 were surveyed by a crew from Colorado State University and cross sections 28, 30, 32, 34, 36 and 38 were surveyed by the USDA, NRCS. Figure 1 shows the path of the low flow channel, the location of each cross section and the location of major large woody debris elements surveyed in June 2000. Cross sections were also surveyed by the US Army Corps of engineers in 1997 but these sections do not tie in with the 1999-2000 locations. A geomorphic evaluation of each cross section was undertaken in June 2000 to assess stage of evolution in the Simon six-stage evolution sequence. Data collected includes channel bed and bankfull widths, berm widths, bank angles, bank materials, processes operating on either bank and the rate of sedimentation/bank failure which was determined using dendrochronological techniques. -

Glacier National Park, 1917
~ ________________ ~'i DEPARTMENT OF THE INTERIOR / FRANKLIN K. LANE. SECRETARY NATIONAL PARK SERVI'CE,/ STEPHEN T. MATHER. DIRECTOR GENERAL ~FO ~N GL CIER NAL ONAL PARK Season of 191 7 The Alps of America-Wonderful Tumbled Region Possessing 60 Glaciers. 250 Lakes, and M y Stately Peaks-Precipices 4,000 Feet Deep-Valleys of Astonish ing Rugged B auty-Scenery Equaling Any in the World- Large, Excellent Hotels and Comfortable Chalet Camps-Good Roads- The Gunsight Trail Across the Top of the Range-Good Trout Fishing-How to Get There-What to See-What to Wear lor MOUiltain Climbing WASHINGTON GOVERNMENT PRINTING OFFICE 1917 TI-IE NATIO .... PARKS AT A GLANCE ( Chron010gIca,l.ly In the order of theIr creatIon [Number,14; Total Area, 7,290 Square Miles] NATIONAL AREA PARKS In DISTINCTIVE CHARACTERISTICS In order of LOCATION square creation miles H ot Springs •..... Middle H 46 hot springs possessing curative properties-Many hotels and 1832 Arkansa.s boording houses-20 bathhouses under public control. CONTENTS. Yellowstone . ••••. North- 3,348 More geysers than in al1 rest of world together-Boiling 1872 western springs-Mud volcanoes-Petrified forests-Grand Canyon Page. Wyoming of the YelIowstone, remarkable for gorgeous coloring-Large General description_ .. _. _. ........ ..... .... ... ... ...... ........ .. 5 lakes-Many large streams and waterfalls-Vast wilderness A romance in rocks . • _. __ . _. _.. .......................... _. ....... 5 inhabited by deer, elk, bison, moose, antelope, bear, moun- The Lewis overthrust .. __. .... _............................... ...... 6 tain sheep, beaver, etc., constituting greatest wild bird and A general view _ . _____ .. ..... ................................. 6 animal preserve in world-Altitude 6,000 to 11,000 feet- The west side .... -

Fluvial Systems – Meandering Rivers Rio Solimoes, Brazil Synthetic Aperture Radar Characteristics of Meandering Rivers
Fluvial systems – meandering rivers Rio Solimoes, Brazil synthetic aperture radar Characteristics of meandering rivers generally confined within one major channel secondary channels active during floods wide valley, channel is a small part of entire valley Characteristics of meandering rivers Compared with braided river: •low gradient • greater sinuosity • greater % suspended load (less bedload) • finer-grained sediments • more constant discharge (usually perennial flow) Meanders, San Joaquin River cohesive banks, little coarse sediment Meanders, Sacramento River (transitional) less cohesive banks, moderate coarse sediment Amazon River meanders an extreme in bank stability (short-term) Scroll plain Rio Apure, Orinoco Basin Meanders and scroll plains Cross section of river valley & channel River valley Active river channel A natural river valley Landforms Note: levees along outside of meanders Meandering and sinuosity Path of highest-velocity flow Point bars lateral accretion of point bars along inside of meander Cut bank and point bar Cut bank, Fountain Creek, New Mexico Point bar, upstream Fountain Creek, New Mexico Point bar, downstream Fountain Creek, New Mexico Flood channel Enhanced turbulence at confluence text Features of a meandering river Figure 5.12a Figure 5.12b Figure 5.12b Figure 5.12c Figure 5.12c Meander cut-off Forming an oxbow lake Overbank deposition Bankfull discharge flood water level up to the top of the channel maintains the primary channel occurs once every 1-2 years Bankfull discharge Bankfull Average flow Figure