End-Of-Day Lighting with Different Red/Far-Red Ratios Using Light
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Iceland Panorama
Iceland Panorama August 5th - 12th 2018 From steamy hot springs to top-notch spas, spectacular scenery to magnificent art museums, this unique land is the perfect place to relax, recharge, and explore. Legends say that the ancient gods themselves guided Iceland’s first settler, Ingolfur Arnarson, to make his home in Reykjavik (“Smoky Bay”), named after the geothermal steam he saw. Today this geothermal energy heats homes and outdoor swimming pools throughout the city – a pollution-free energy source that leaves the air outstandingly fresh, clean and clear. Renowned for its spellbinding beauty and stunning landscape, Iceland is the only place on Earth where you can stand on top of the Atlantic Ocean’s submarine mountain chain - for the island is the only place where the chain peaks up above sea level! From this perch, we can walk from North America to Europe. We’ll journey through the north, south, and west of the island, enjoying the long evening twilight of the northern summer and visit a wondrous variety of small settlements, good-size towns and amazing natural landscapes. This captivating geological wonderland offers the opportunity to explore dramatic phenomena: colossal glaciers, active volcanoes, geysers, hot springs, glacial rivers, cascading waterfalls, moss-covered lava fields, and glacial lagoons. We’ll dine on Icelandic specialties, including seafood, ocean-fresh from the morning’s catch; highland lamb; and unusual varieties of game. It’s purely natural food imaginatively served. The cost of this itinerary, per person, double occupancy is: Join a Tufts faculty host and specially selected local guide, Boston Departure $3990 Svanur Thorkelsson on an amazing adventure this summer Land only (no airfare included) $3290 to the Land of Fire and Ice. -
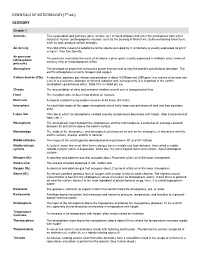
ESSENTIALS of METEOROLOGY (7Th Ed.) GLOSSARY
ESSENTIALS OF METEOROLOGY (7th ed.) GLOSSARY Chapter 1 Aerosols Tiny suspended solid particles (dust, smoke, etc.) or liquid droplets that enter the atmosphere from either natural or human (anthropogenic) sources, such as the burning of fossil fuels. Sulfur-containing fossil fuels, such as coal, produce sulfate aerosols. Air density The ratio of the mass of a substance to the volume occupied by it. Air density is usually expressed as g/cm3 or kg/m3. Also See Density. Air pressure The pressure exerted by the mass of air above a given point, usually expressed in millibars (mb), inches of (atmospheric mercury (Hg) or in hectopascals (hPa). pressure) Atmosphere The envelope of gases that surround a planet and are held to it by the planet's gravitational attraction. The earth's atmosphere is mainly nitrogen and oxygen. Carbon dioxide (CO2) A colorless, odorless gas whose concentration is about 0.039 percent (390 ppm) in a volume of air near sea level. It is a selective absorber of infrared radiation and, consequently, it is important in the earth's atmospheric greenhouse effect. Solid CO2 is called dry ice. Climate The accumulation of daily and seasonal weather events over a long period of time. Front The transition zone between two distinct air masses. Hurricane A tropical cyclone having winds in excess of 64 knots (74 mi/hr). Ionosphere An electrified region of the upper atmosphere where fairly large concentrations of ions and free electrons exist. Lapse rate The rate at which an atmospheric variable (usually temperature) decreases with height. (See Environmental lapse rate.) Mesosphere The atmospheric layer between the stratosphere and the thermosphere. -

Romancing the Stockholm Syndrome: Desensitizing Readers to Obsessive and Abusive Behavior in Contemporary Young Adult Literature
Romancing the Stockholm Syndrome: Desensitizing Readers to Obsessive and Abusive Behavior in Contemporary Young Adult Literature Lauren Staub Honors Thesis, Spring 2013 Thesis Director: Terry Harpold Reader: Anastasia Ulanowicz Contemporary works of young adult (or “YA”) literature written and marketed primarily for adolescent girls often depend upon conventional narratives that feature the damsel in distress – that is, plots in which the young woman who needs to be saved from her surroundings, or even herself, by a young man. Such privileging of masculine power – even in contemporary and purportedly progressive narratives – reaffirms the conservative notion that men still hold the power in relationships, that men's mastery over their female partners is an appropriate expression of passion and attraction. But what happens when the characters in such texts – as well as their readers – confuse abuse for passion? Young women reading stories that portray controlling or over-protective men as paradigms of romance are, I propose, being desensitized to obsessive and abusive behavior. Forced captivity and stalking, along with other dangerous behaviors should not be mistaken for passionate love. Passive surrender to overheated, manipulative and aggressive suitors (the classic "bad boy" of these YA novels) means surrendering independent agency and defining desire and love in only reactive ways. This is not a positive model for female readers. Page 1 of 26 In this thesis, then, I will analyze three popular YA novels written primarily for female readers in order to study how the novels employ, and occasionally justify kidnapping and Stockholm Syndrome plots. I will briefly consider literature on the psychiatric condition of Stockholm Syndrome and its popular representation in fiction. -

Summer Solstice
Summer Solstice The equator is an imaginary line around the middle of the Earth. Above the equator is the northern hemisphere. Below the equator is the southern hemisphere. Can you imagine a pole going through Earth from the North Pole to the South Pole? This pole would be the Earth’s axis. The Earth spins around this axis. The axis is not vertical; it tilts the Earth over. This means the Earth appears to lean over. The Earth orbits or moves on a path around the Sun. This takes around one year. At different times of the year, as it journeys around the Sun, some places on Earth are nearer to the Sun than others. If you live above the equator, Earth is tilted closer to the Sun in the summer, giving more light and heat. In winter, these countries are further away from the sun and have less light and heat. Spring Sun Winter Summer Autumn This diagram shows the seasons in the northern hemisphere, above the equator. Can you see how the northern hemisphere is tilted towards the sun in summer? Page 1 of 3 Summer Solstice What is the Summer Solstice? The Summer Solstice happens when the North Pole is most tilted towards the sun. It marks the change when days in the northern hemisphere begin to grow shorter. The Summer Solstice happens around 21st June. This is also known as midsummer and is the longest day and shortest night of the year in the northern hemisphere. Summer Solstice in the Far North Solstice Celebrations Around the Summer Solstice, countries For thousands of years, there have in the Arctic Circle, like parts of been solstice celebrations around Norway, Finland, Greenland and the world. -

CHAPTER I INTRODUCTION A. Background of the Study Twilight Is One of the Most Popular Movies of This Year That Ever Produced. Th
1 CHAPTER I INTRODUCTION A. Background of the Study Twilight is one of the most popular movies of this year that ever produced. The screenplay of this film is adapted from the novel with the same title written by Stephenie Meyer. Stephenie Meyer is an American author best known for her vampire romance series Twilight who was born on December 24, 1973 in Hartford. The Twilight novels have gained worldwide recognition, won multiple literary awards and sold over 85million copies worldwide, with translation into 37 different languages. Meyer is also the author of the adult science-fiction novel The Host (2008). Following the success of Twilight (2005), Meyer expended the story into a series three more books: New Moon (2006), Eclipse (2007), and Breaking Down (2008). She also writes short stories which one of them published in Prom Nights from Hell, a collection stories about bad prom nights with supernatural effects. It has been released in April 2007. Twilight movie is directed by Catherine Hardwicke, an American production designer and film director and screenplay by Melissa Rosenberg. Twilight is better known as the maker romance and fantasy film, sometimes centering on vampire society’s life. Twilight is a 2008 romantic fantasy film. It is the first film in The Twilight saga film series. Twilight focuses on the development of a personal relationship between human teenager Bella Swan and vampire Edward Cullen and the subsequent efforts of Cullen and his family to keep Swan safe from a separate group of hostile vampires. Seventeen-year-old Isabella "Bella" Swan moves to Forks, a small town near Washington State’s rugged coast, to live with her father, Charlie, after her mother remarries to a minor league baseball player. -

Intertextuality, Masculinity, and Desire in the Twilight Series
Virtue as Adventure and Excess: Intertextuality, Masculinity, and Desire in the Twilight Series By Claudia Lindén Abstract The vampire is still primarily a literary figure. The vampires we have seen on TV and cinema in recent years are all based on literary models. The vampire is at the same time a popular cultural icon and a figure that, especially women writers, use to problematize gender, sexuality and power. As a vampire story the Twilight se- ries both produces and problematizes norms in regard to gender, class and ethnici- ty. As the main romantic character in Twilight, Edward Cullen becomes interest- ing both as a vampire of our time and as a man. In a similar way as in the 19th century novel the terms of relationship are negotiated and like his namesake Ed- ward Rochester, Edward Cullen has to change in important ways for the “happy ending” to take place. In spite of a strong interest in sexuality and gender norms in relation to vampires very few studies have focused exclusively on masculinity. This article examines the construction of masculinity in relation to vampirism in the Twilight series. It offers an interpretation of Stephenie Meyer’s novels and the character of Edward as part of a broader field of feminist (re-)uses of the vampire in modern literature with its roots in the literary tradition from Austen and the Brontë-sisters as well as from classic Gothic fiction. Keywords: Twilight Series, Stephenie Meyer, masculinity, vampires, were- wolves, Midnight Sun, Jane Austen, Pride and Prejudice, Charlotte Brontë, Jane Eyre, feminist theory, queer theory, gothic. -

Planit! User Guide
ALL-IN-ONE PLANNING APP FOR LANDSCAPE PHOTOGRAPHERS QUICK USER GUIDES The Sun and the Moon Rise and Set The Rise and Set page shows the 1 time of the sunrise, sunset, moonrise, and moonset on a day as A sunrise always happens before a The azimuth of the Sun or the well as their azimuth. Moon is shown as thick color sunset on the same day. However, on lines on the map . some days, the moonset could take place before the moonrise within the Confused about which line same day. On those days, we might 3 means what? Just look at the show either the next day’s moonset or colors of the icons and lines. the previous day’s moonrise Within the app, everything depending on the current time. In any related to the Sun is in orange. case, the left one is always moonrise Everything related to the Moon and the right one is always moonset. is in blue. Sunrise: a lighter orange Sunset: a darker orange Moonrise: a lighter blue 2 Moonset: a darker blue 4 You may see a little superscript “+1” or “1-” to some of the moonrise or moonset times. The “+1” or “1-” sign means the event happens on the next day or the previous day, respectively. Perpetual Day and Perpetual Night This is a very short day ( If further north, there is no Sometimes there is no sunrise only 2 hours) in Iceland. sunrise or sunset. or sunset for a given day. It is called the perpetual day when the Sun never sets, or perpetual night when the Sun never rises. -

June Solstice Activities (PDF)
Arctic Connection Linking Your Place to the MOSAiC Expedition June Solstice Edition Introduction As I write this, it is the June Solstice. The exact moment of solstice occurred a few hours ago, at 21:44 Universal Daylight Time. This was at 1:44 pm today here in Homer, Alaska. This moment marked when Earth’s north pole leaned most toward the sun, and the Earth’s south pole was tilted most away from the sun. On this day, the sun appears directly overhead at local noon for those living at 23.5 degrees north (the Tropic of Cancer), as far north as the sun ever gets. And during the December solstice, the sun appears directly overhead for those living at 23.5 degrees south (the Tropic of Capricorn). (In case you need a refresher, here’s the basic science from Earth & Sky.) In the northern hemisphere, the June Solstice is called the summer solstice and represents the day(s) with the most amount of daylight. I say days because in some parts of the northern hemisphere, the sun has stayed above the horizon for multiple days now and won’t rise again until the next month. This is often called the Midnight Sun, and the ice camp at the Polarstern has been bathed in light for many days. This is good news for the scientists of Leg 4, who are just now arriving to the floe. The extended daylight will help make all of the research tasks a little bit easier than those Leg 1 and Leg 2 researchers who had to work through the Polar Night. -

Understanding Golden Hour, Blue Hour and Twilights
Understanding Golden Hour, Blue Hour and Twilights www.photopills.com Mark Gee proves everyone can take contagious images 1 Feel free to share this ebook © PhotoPills April 2017 Never Stop Learning The Definitive Guide to Shooting Hypnotic Star Trails How To Shoot Truly Contagious Milky Way Pictures A Guide to the Best Meteor Showers in 2017: When, Where and How to Shoot Them 7 Tips to Make the Next Supermoon Shine in Your Photos MORE TUTORIALS AT PHOTOPILLS.COM/ACADEMY Understanding How To Plan the Azimuth and Milky Way Using Elevation The Augmented Reality How to find How To Plan The moonrises and Next Full Moon moonsets PhotoPills Awards Get your photos featured and win $6,600 in cash prizes Learn more+ Join PhotoPillers from around the world for a 7 fun-filled days of learning and adventure in the island of light! Learn More We all know that light is the crucial element in photography. Understanding how it behaves and the factors that influence it is mandatory. For sunlight, we can distinguish the following light phases depending on the elevation of the sun: golden hour, blue hour, twilights, daytime and nighttime. Starting time and duration of these light phases depend on the location you are. This is why it is so important to thoughtfully plan for a right timing when your travel abroad. Predicting them is compulsory in travel photography. Also, by knowing when each phase occurs and its light conditions, you will be able to assess what type of photography will be most suitable for each moment. Understanding Golden Hour, Blue Hour and Twilights 6 “In almost all photography it’s the quality of light that makes or breaks the shot. -

Terminology Guidelines for the Protection of Children from Sexual Exploitation and Sexual Abuse
TERMINOLOGY GUIDELINES FOR THE PROTECTION OF CHILDREN FROM SEXUAL EXPLOITATION AND SEXUAL ABUSE Terminology Guidelines for the Protection of Children from Sexual Exploitation and Sexual Abuse Adopted by the Interagency Working Group in Luxembourg, 28 January 2016 Text written by Susanna Greijer and Jaap Doek and approved by the Interagency Working Group ISBN: 978-92-61-21491-3 (paper version) 978-92-61-21501-9 (electronic version) English original, first printing, June 2016 June, 2016 Copyright ECPAT International, jointly with ECPAT Luxembourg This work is licensed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License. To view a copy of this license, visit http:// creativecommons. org/ licenses/by- nc- nd/ 4. 0/ . Permission to translate this work should be obtained through ECPAT International 328/1 Phayathai Road Rachathewi, Bangkok 10400 Thailand Tel: +66 2 215 3388 Fax: +66 2 215 8272 Email: [email protected] Website: www. ecpat.net Acknowledgements The content of this publication is shaped from discussions by the Interagency Working Group (IWG) and input from the participating organisations, and particular thanks are expressed to the following persons: Maud de Boer-Buquicchio (UN Special Rapporteur on the sale of children, child prostitution and child pornography) and relevant staff members from the Office of the UN High Commissioner for Human Rights, Sheila Varadan, ECPAT International, Benyam Mezmur (African Committee on the Rights and Welfare of the Child), Beatrice Schulter and Anita Goh (Child -

Printed Fabric Brochure Contents…
Printed Fabric Brochure Contents… 3 Printed Fabric Collections 4 Printed Semi Plain Designs 5 Melrose Collection 8 Mayfield Collection 10 Nostalgia Collection 12 Chatsworth Collection 14 Copenhagen Collection 16 Wordsworth Collection 18 Capture Collection 20 Seasons Collection 22 Botanical Collection 24 Novello Collection 25 Ophelia Collection 26 Aurora Plain Bedding (Q1739) Printed Fabric Collections… 3 Twilight Twilight Natural Twilight Twilight Straw Twilight Camel Twilight Latte Twilight Taupe Twilight Almond Hessian Oatmeal PLAIN DESIGNS PLAIN Twilight Misty Twilight Mink Twilight Peat Twilight Twilight Twilight Ice Twilight Silver Twilight Steel Cappuccino Platinum PRINTED SEMI SEMI PRINTED Twilight Smoke Twilight Slate Twilight Pewter Twilight Twilight Bark Twilight Noir Charcoal Twilight Citrus Twilight Twilight Saffron Twilight Soft Twilight Willow Twilight Apple Twilight Khaki Twilight Pine Sunshine Green Twilight Twilight Spa Twilight Sky Twilight Cobalt Twilight Cyan Twilight Teal Twilight Twilight Eau de Nil Emerald Midnight Twilight Blush Twilight Twilight Rose Twilight Coral Twilight Twilight Papaya Twilight Spice Twilight Damson Crimson Bordeaux Twilight Twilight Plum Twilight Twilight Twilight Navy Heather Mulberry Aubergine All of the designs throughout this brochure can be printed on to any of our 17 base cloths, making them suitable for a variety of end uses from waterproof upholstery to satin curtaining. 4 Helbeck Ivory Helbeck Smoke Helbeck Putty Helbeck Sand Helbeck Helbeck Helbeck Helbeck Noir Hessian Charcoal Shadow -

St. Petersburg: “Marooned on the Neva's Marsh Delta”
Giblett, Rod. "St. Petersburg: “Marooned on the Neva’s Marsh Delta”." Cities and Wetlands: The Return of the Repressed in Nature and Culture. London: Bloomsbury Academic, 2016. 125–142. Environmental Cultures. Bloomsbury Collections. Web. 28 Sep. 2021. <http:// dx.doi.org/10.5040/9781474269858.ch-008>. Downloaded from Bloomsbury Collections, www.bloomsburycollections.com, 28 September 2021, 21:45 UTC. Copyright © Rod Giblett 2016. You may share this work for non-commercial purposes only, provided you give attribution to the copyright holder and the publisher, and provide a link to the Creative Commons licence. 8 St. Petersburg: “Marooned on the Neva’s Marsh Delta” St. Petersburg is another marsh metropolis and an icon of modernity. St. Petersburg for Amery and Curran (2006: 15) is “one of the most celebrated and legendary cities in all of Europe.” Certainly it is one of the most cel- ebrated and legendary cities in modern Europe, and is arguably the mod- ern European city par excellence as most of the other major European cities had medieval or ancient beginnings as we have seen with Paris, Venice, and London. Founded in 1703, it achieved this status precisely because it was “situated unnaturally in a swamp,” or “on inhospitable marshes,” or “over swampland” as Lehan (1998: 125, 146, 147) variously puts it. St. Petersburg represents the triumph of a modern city over premodern wetland. Like Venice and Boston, St. Petersburg was built on piles. Even with its piles, Th oreau (1982: 276) relates in Walden how “it is said that a fl ood tide, with a westerly wind and ice in the Neva, would sweep St.