Ewa Żurawska-Seta
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Revision of the Mole Genus Mogera (Mammalia: Lipotyphla: Talpidae)
Systematics and Biodiversity 5 (2): 223–240 Issued 25 May 2007 doi:10.1017/S1477200006002271 Printed in the United Kingdom C The Natural History Museum Shin-ichiro Kawada1, 6 , Akio Shinohara2 , Shuji Revision of the mole genus Mogera Kobayashi3 , Masashi Harada4 ,Sen-ichiOda1 & (Mammalia: Lipotyphla: Talpidae) Liang-Kong Lin5 ∗ 1Laboratory of Animal from Taiwan Management and Resources, Graduate School of Bio-Agricultural Sciences, Nagoya University, Nagoya, Aichi 464-8601, Japan Abstract We surveyed the central mountains and southeastern region of Taiwan 2Department of Bio-resources, and collected 11 specimens of a new species of mole, genus Mogera. The specimens Division of Biotechnology, were characterized by a small body size, dark fur, a protruding snout, and a long Frontier Science Research Center, University of Miyazaki tail; these characteristics are distinct from those of the Taiwanese lowland mole, 5200, Kihara, Kiyotake, M. insularis (Swinhoe, 1862). A phylogenetic study of morphological, karyological Miyazaki 889-1692, Japan 3Department of Asian Studies, and molecular characters revealed that Taiwanese moles should be classified as two Chukyo Woman’s University, distinctive species: M. insularis from the northern and western lowlands and the new Yokone-cho, Aichi 474-0011, species from the central mountains and the east and south of Taiwan. The skull of Japan 4Laboratory Animal Center, the new species was slender and delicate compared to that of M. insularis. Although Osaka City University Graduate the karyotypes of two species were identical, the genetic distance between them School of Medical School, Osaka, Osaka 545-8585, Japan was sufficient to justify considering each as a separate species. Here, we present a 5Laboratory of Wildlife Ecology, detailed specific description of the new species and discuss the relationship between Department of Life Science, this species and M. -

Controlled Animals
Environment and Sustainable Resource Development Fish and Wildlife Policy Division Controlled Animals Wildlife Regulation, Schedule 5, Part 1-4: Controlled Animals Subject to the Wildlife Act, a person must not be in possession of a wildlife or controlled animal unless authorized by a permit to do so, the animal was lawfully acquired, was lawfully exported from a jurisdiction outside of Alberta and was lawfully imported into Alberta. NOTES: 1 Animals listed in this Schedule, as a general rule, are described in the left hand column by reference to common or descriptive names and in the right hand column by reference to scientific names. But, in the event of any conflict as to the kind of animals that are listed, a scientific name in the right hand column prevails over the corresponding common or descriptive name in the left hand column. 2 Also included in this Schedule is any animal that is the hybrid offspring resulting from the crossing, whether before or after the commencement of this Schedule, of 2 animals at least one of which is or was an animal of a kind that is a controlled animal by virtue of this Schedule. 3 This Schedule excludes all wildlife animals, and therefore if a wildlife animal would, but for this Note, be included in this Schedule, it is hereby excluded from being a controlled animal. Part 1 Mammals (Class Mammalia) 1. AMERICAN OPOSSUMS (Family Didelphidae) Virginia Opossum Didelphis virginiana 2. SHREWS (Family Soricidae) Long-tailed Shrews Genus Sorex Arboreal Brown-toothed Shrew Episoriculus macrurus North American Least Shrew Cryptotis parva Old World Water Shrews Genus Neomys Ussuri White-toothed Shrew Crocidura lasiura Greater White-toothed Shrew Crocidura russula Siberian Shrew Crocidura sibirica Piebald Shrew Diplomesodon pulchellum 3. -
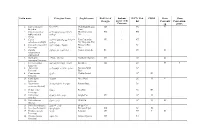
Red List of Endemic IUCN Red CITES Bern Bonn Georgia Species of the List Conventi Convention Caucasus on (CMS) 1
Latin name Georgian Name English name Red List of Endemic IUCN Red CITES Bern Bonn Georgia species of the list Conventi Convention Caucasus on (CMS) 1. Capra aegagrus niamori Wild Goat, Bezoar CR VU II Erxleben. Goat 2. Capra caucasica dasavleTkavkasiuri West Caucasian EN + EN Güldenstädt & jixvi Tur Pallas. 3. Capra aRmosavleTkavkasiuri East Caucasian VU + NT cylindricornis Blyth. jixvi Tur, Dagestan Tur 4. Capreolus capreolus evropuli Sveli European Roe LC Linnaeus. Deer 5. Gazella qurciki, jeirani Goitered Gazelle RE VU II subgutturosa Güldenstädt. 6. Rupicapra arCvi, fsiti Northern Chamois EN LC II rupicapra Linnaeus. 7. Cervus elaphus keTilSobili iremi Red Deer CR LC II I Linnaeus. 8. Sus scrofa gareuli Rori, taxi Eurasian Wild LC Linnaeus. Boar 9. Canis aureus tura Golden Jackal LC III Linnaeus. 10. Canis lupus mgeli Grey Wolf LC II II Linnaeus. 11. Nyctereutes enotisebri ZaRli Racoon Dog LC procyonoides Gray. 12. Vulpes vulpes mela Red Fox LC III Linnaeus. 13. Felis chaus lelianis kata Jungle Cat VU LC II Schreber. 14. Felis silvestris tyis kata Wild Cat LC II II Shreber. 15. Felis libyca Forster. velis kata Steppe Cat 16. Lynx lynx Linnaeus. focxveri Eurasian Lynx CR LC II 17. Panthera pardus jiqi Leopard CR NT I II Linnaeus. 18. Hyaena hyaena afTari Striped Hyaena CR NT Linnaeus. 19. Lutra lutra wavi Eurasian Otter, VU NT I II Linnaeus. Common Otter 20. Martes foina kldis kverna Stone Marten, LC III Erxleben. Beech Marten 21. Martes martes tyis kverna European Pine LC Linnaeus. Marten 22. Meles meles maCvi Eurasian Badger LC Linnaeus. 23. Mustela lutreola waula European Mink EN II Linnaeus. -

Comparative Morphology of the Penis and Clitoris in Four Species of Moles
RESEARCH ARTICLE Comparative Morphology of the Penis and Clitoris in Four Species of Moles (Talpidae) ADRIANE WATKINS SINCLAIR1∗, STEPHEN GLICKMAN2, KENNETH CATANIA3, AKIO SHINOHARA4, LAWRENCE BASKIN1, 1 AND GERALD R. CUNHA 1Department of Urology, University of California San Francisco, San Francisco, California 2Departments of Psychology and Integrative Biology, University of California, Berkeley, California 3Department of Biological Sciences, Vanderbilt University, Nashville, Tennessee 4Frontier Science Research Center, University of Miyazaki, Kihara, Japan ABSTRACT The penile and clitoral anatomy of four species of Talpid moles (broad-footed, star-nosed, hairy- tailed, and Japanese shrew moles) were investigated to define penile and clitoral anatomy and to examine the relationship of the clitoral anatomy with the presence or absence of ovotestes. The ovotestis contains ovarian tissue and glandular tissue resembling fetal testicular tissue and can produce androgens. The ovotestis is present in star-nosed and hairy-tailed moles, but not in broad-footed and Japanese shrew moles. Using histology, three-dimensional reconstruction, and morphometric analysis, sexual dimorphism was examined with regard to a nine feature mascu- line trait score that included perineal appendage length (prepuce), anogenital distance, and pres- ence/absence of bone. The presence/absence of ovotestes was discordant in all four mole species for sex differentiation features. For many sex differentiation features, discordance with ovotestes was observed in at least one mole species. The degree of concordance with ovotestes was highest for hairy-tailed moles and lowest for broad-footed moles. In relationship to phylogenetic clade, sex differentiation features also did not correlate with the similarity/divergence of the features and presence/absence of ovotestes. -

An Evolutionary View on the Japanese Talpids Based on Nucleotide Sequences
Mammal Study 30: S19–S24 (2005) © the Mammalogical Society of Japan An evolutionary view on the Japanese talpids based on nucleotide sequences Akio Shinohara1,*, Kevin L. Campbell2 and Hitoshi Suzuki3 1 Department of Bio-resources, Division of Biotechnology, Frontier Science Research Center, University of Miyazaki, Miyazaki 889-1692, Japan 2 Department of Zoology, University of Manitoba, Winnipeg, Manitoba, R3T 2N2, Canada 3 Graduate School of Environmental Earth Science, Hokkaido University, Hokkaido 060-0810, Japan Abstract. Japanese talpid moles exhibit a remarkable degree of species richness and geographic complexity, and as such, have attracted much research interest by morphologists, cytogeneticists, and molecular phylogeneticists. However, a consensus hypothesis pertaining to the evolutionary history and biogeography of this group remains elusive. Recent phylogenetic studies utilizing nucleotide sequences have provided reasonably consistent branching patterns for Japanese talpids, but have generally suffered from a lack of closely related South-East Asian species for sound biogeographic interpretations. As an initial step in achieving this goal, we constructed phylogenetic trees using publicly accessible mitochondrial and nuclear sequences from seven Japanese taxa, and those of related insular and continental species for which nucleotide data is available. The resultant trees support the view that four lineages (Euroscaptor mizura, Mogera tokuade species group [M. tokudae and M. etigo], M. imaizumii, and M. wogura) migrated separately, and in this order, from the continental Asian mainland to Japan. The close relationship of M. tokudae and M. etigo suggests these lineages diverged recently through a vicariant event between Sado Island and Echigo plain. The origin of the two endemic lineages of Japanese shrew-moles, Urotrichus talpoides and Dymecodon pilirostris, remains ambiguous. -

Activity of the European Mole Talpa Europaea (Talpidae, Insectivora) in Its Burrows in the Republic of Mordovia
FORESTRY IDEAS, 2021, vol. 27, No 1 (61): 59–67 ACTIVITY OF THE EUROPEAN MOLE TALPA EUROPAEA (TALPIDAE, INSECTIVORA) IN ITS BURROWS IN THE REPUBLIC OF MORDOVIA Alexey Andreychev Department of Zoology, National Research Mordovia State University, Saransk 430005, Russia. E-mail: [email protected] Received: 16 February 2021 Accepted: 18 April 2021 Abstract A new method of studying the activity of European mole Talpa europaea (Linnaeus, 1758) with use of digital portable voice recorders is developed. European mole demonstrates polyphasic activity pattern – three peaks of activity alternate three peaks of relative rest. Moles are found to have activity peaks from 23:00 to 3:00 h, from 6:00 to 9:00 h and from 15:00 to 18:00 h, and three periods of rest: from 3:00 to 6:00 h, from 9:00 to 15:00 h and from 18:00 to 23:00 h. Mole’s rest is relative since animals show low activity during periods of rest. Average daily interval between mole passes is 2.5 h. Duration of audibility of continuous single European mole pass by the mi- crophone varies from 11 to 120 seconds. On average, it is 37.5 seconds. Key words: animals, daily activity, day-night activity, voice recorder. Introduction where light factor is essential (Shilov 2001). Activity of many mammals is in- In many countries, the European mole Tal- vestigated under various conditions of the pa europaea (Linnaeus, 1758) is an object light regime. For underground animals, of constant modern research (Komarnicki especially for the different mole species, 2000, Prochel 2006, Kang et al. -

What Should We Call the Levant Mole? Unravelling the Systematics and Demography of Talpa Levantis Thomas, 1906 Sensu Lato (Mammalia: Talpidae)
University of Plymouth PEARL https://pearl.plymouth.ac.uk Faculty of Science and Engineering School of Biological and Marine Sciences 2020-03-02 What should we call the Levant mole? Unravelling the systematics and demography of Talpa levantis Thomas, 1906 sensu lato (Mammalia: Talpidae) Demirtas, S http://hdl.handle.net/10026.1/15424 10.1007/s42991-020-00010-4 Mammalian Biology Elsevier All content in PEARL is protected by copyright law. Author manuscripts are made available in accordance with publisher policies. Please cite only the published version using the details provided on the item record or document. In the absence of an open licence (e.g. Creative Commons), permissions for further reuse of content should be sought from the publisher or author. 1 What should we call the Levant mole? Unravelling the systematics and demography of 2 Talpa levantis Thomas, 1906 sensu lato (Mammalia: Talpidae). 3 4 Sadik Demirtaşa, Metin Silsüpüra, Jeremy B. Searleb, David Biltonc,d, İslam Gündüza,* 5 6 aDepartment of Biology, Faculty of Arts and Sciences, Ondokuz Mayis University, Samsun, 7 Turkey. 8 bDepartment of Ecology and Evolutionary Biology, Cornell University, Ithaca, NY, 14853- 9 2701, USA. 10 cMarine Biology and Ecology Research Centre, School of Biological and Marine Sciences, 11 University of Plymouth, Plymouth PL4 8AA, Devon, UK. 12 dDepartment of Zoology, University of Johannesburg, PO Box 524, Auckland Park, 13 Johannesburg 2006, Republic of South Africa 14 15 *Corresponding author. E-mail: [email protected] 16 1 17 Abstract 18 19 Turkey hosts five of the eleven species of Talpa described to date, Anatolia in particular 20 appearing to be an important centre of diversity for this genus. -

Cytogenetic Peculiarities in the Algerian Hedgehog: Silver Stains Not Only Nors but Also Heterochromatic Blocks
Heredity 75 (1995)10-16 Received24 August 1994 Cytogenetic peculiarities in the Algerian hedgehog: silver stains not only NORs but also heterochromatic blocks ANTONIO SANCHEZ, RAFAEL JIMENEZ*, MIGUEL BURGOS, SAIDA STITOU, FEDERICO ZURITA & RAFAEL D1AZ DE LA GUARDIA Departamento de Genética, Facu/tad de Ciencias, Universidad de Granada, 18071 Granada, Spain Hedgehogsbelong to one of the several mammalian taxa in which karyotype differences are based on variations in heterochromatin content. Furthermore, the number and location of nucleolar organizer regions (NORs) can also vary widely. In the present study these cytogenetic features were investigated in the Algerian hedgehog, Erinaceus (Aethechinus) algirus. The heterochromatin and NOR distribution patterns in the karyotype of this species are new among hedgehogs, whereas the euchromatic regions, including their G-band pattern, are similar to those reported by others. In addition, silver staining revealed a cytogenetic feature exclusive to the heterochromatic blocks of E. algirus: their silver staining with standard cytogenetic procedures. Because no similar phenomenon has been described previously in a mammalian species, several hypotheses about the significance and specificity of silver staining to NOR sites are discussed. Finally, the existence of different types of heterochromatin in the species analysed here, lead us to propose that what hedgehogs have inherited from their common ancestor is a mechanism which permits the accumulation of hetero- chromatin on specific chromosomes, rather than the heterochromatin itself. Keywords: Erinaceusalgirus, heterochromatinvariation, karyotype evolution, NOR variation, silver staining. Introduction blocks. In this species, however, their relationship with NORs could not be established, as Ag-staining tech- Hedgehogshave been the subject of several interesting niques were not available when this work was done. -

INSIGHTS INTO RELATIONSHIPS AMONG RODENT LINEAGES BASED on MITOCHONDRIAL GENOME SEQUENCE DATA a Dissertation by LAURENCE JOHN FR
INSIGHTS INTO RELATIONSHIPS AMONG RODENT LINEAGES BASED ON MITOCHONDRIAL GENOME SEQUENCE DATA A Dissertation by LAURENCE JOHN FRABOTTA Submitted to the Office of Graduate Studies of Texas A&M University in partial fulfillment of the requirements for the degree of DOCTOR OF PHILOSOPHY December 2005 Major Subject: Zoology INSIGHTS INTO RELATIONSHIPS AMONG RODENT LINEAGES BASED ON MITOCHONDRIAL GENOME SEQUENCE DATA A Dissertation by LAURENCE JOHN FRABOTTA Submitted to the Office of Graduate Studies of Texas A&M University in partial fulfillment of the requirements for the degree of DOCTOR OF PHILOSOPHY Approved by: Chair of Committee, Rodney L. Honeycutt Committee Members, James B. Woolley John W. Bickham James R. Manhart Head of Department, Vincent M. Cassone December 2005 Major Subject: Zoology iii ABSTRACT Insights into Relationships among Rodent Lineages Based on Mitochondrial Genome Sequence Data. (December 2005) Laurence John Frabotta, B.S.; M.S., California State University, Long Beach Chair of Advisory Committee: Dr. Rodney L. Honeycutt This dissertation has two major sections. In Chapter II, complete mitochondrial (mt DNA) genome sequences were used to construct a hypothesis for affinities of most major lineages of rodents that arose quickly in the Eocene and were well established by the end of the Oligocene. Determining the relationships among extant members of such old lineages can be difficult. Two traditional schemes on subordinal classification of rodents have persisted for over a century, dividing rodents into either two or three suborders, with relationships among families or superfamilies remaining problematic. The mtDNA sequences for four new rodent taxa (Aplodontia, Cratogeomys, Erethizon, and Hystrix), along with previously published Euarchontoglires taxa, were analyzed under parsimony, likelihood, and Bayesian criteria. -

Guide to Nature Tourism in the Algarve Contents Preface
guide to nature tourism in the algarve Contents Preface 2 Introduction Algarve’s (still) hidden natural treasures 5 Algarve - brief ecogeographic characterization 9 Costa Vicentina The Algarve is naturally big. Or rather big, naturally. This territory of nearly five thousand square 14 Planalto Vicentino kilometres goes far beyond the sands that concentrate the attention of tourists in the summer. 20 Paleodunes From the Barlavento to the Sotavento, there are hidden protected natural areas or that haven’t 23 Reserva Biogenética de Sagres had the attention they deserve. They all wait patiently in the wild, for the looks and the passing 28 Estuaries and Marine Environment by of true Nature lovers. If you are one of them, I warn you that here, in these pages, the journey 33 South Coast begins, through the cliffs, the forest, the riparian corridors and estuarine systems of the Algarve. 38 Costal Wetlands 40 Ria Formosa In fact, we have so many landscapes and with such great biological importance that choosing 47 Ria de Alvor just one would be a Herculean task. And if there is no possible selection among the various 51 Estuário do Arade natural environments in the Algarve, the only solution is to discover them all. Slowly and with 54 Marshes, Reedbeds and Coastal Lagoons heightened senses. 59 Dune Systems and Pine Forests 65 Cliffs at Central Algarve This guide therefore strengthens the diversity of experiences that the destination offers to those 69 Barrocal who want to turn relaxation time into a real vacation. It is no coincidence that nature tourism 76 Fonte da Benémola emerges as one of the products that are “in development” in the Algarve, in the revision proposal 79 Ribeira de Quarteira document of the National Strategic Plan for Tourism (PENT). -

Terrestrial Ecology
Chapter 11: Terrestrial Ecology URS-EIA-REP-204635 Table of Contents 11 Terrestrial Ecology ................................................................................... 11-1 11.1 Introduction ...................................................................................................... 11-1 11.2 Scoping ............................................................................................................ 11-1 11.2.1 ENVIID ................................................................................................ 11-2 11.2.2 Stakeholder Engagement ...................................................................... 11-2 11.2.3 Analysis of Alternatives ......................................................................... 11-4 11.3 Spatial and Temporal Boundaries ........................................................................ 11-4 11.3.1 Spatial Boundaries ................................................................................ 11-4 11.3.2 Temporal Boundaries .......................................................................... 11-11 11.4 Baseline Data .................................................................................................. 11-11 11.4.1 Introduction ....................................................................................... 11-11 11.4.2 Secondary Data .................................................................................. 11-11 11.4.3 Data Gaps .......................................................................................... 11-14 -

Coccidia (Apicomplexa: Eimeriidae) of the Mammalian Order Insectivora
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Faculty Publications from the Harold W. Manter Laboratory of Parasitology Parasitology, Harold W. Manter Laboratory of 2000 Coccidia (Apicomplexa: Eimeriidae) of the Mammalian Order Insectivora Donald W. Duszynski University of New Mexico, [email protected] Steve J. Upton Kansas State University Follow this and additional works at: https://digitalcommons.unl.edu/parasitologyfacpubs Part of the Parasitology Commons Duszynski, Donald W. and Upton, Steve J., "Coccidia (Apicomplexa: Eimeriidae) of the Mammalian Order Insectivora" (2000). Faculty Publications from the Harold W. Manter Laboratory of Parasitology. 196. https://digitalcommons.unl.edu/parasitologyfacpubs/196 This Article is brought to you for free and open access by the Parasitology, Harold W. Manter Laboratory of at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Faculty Publications from the Harold W. Manter Laboratory of Parasitology by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. SPECIAL PUBLICATION THE MUSEUM OF SOUTHWESTERN BIOLOGY NUMBER 4, pp. 1-67 30 OCTOBER 2000 Coccidia (Apicomplexa: Eimeriidae) of the Mammalian Order Insectivora DONALD W. DUSZYNSKI AND STEVE J. UPTON TABLE OF CONTENTS Introduction 1 Materials and Methods 2 Results 3 Family Erinaceidae Erinaceus Eimeria ostertagi 3 E. perardi 4 Isospora erinacei 4 I. rastegaievae 5 I. schmaltzi 6 Hemiechinus E. auriti 7 E. bijlikuli 7 Hylomys E. bentongi 7 I. hylomysis 8 Family Soricidae Crocidura E. firestonei 8 E. leucodontis 9 E. milleri 9 E. ropotamae 10 Suncus E. darjeelingensis 10 E. murinus...................................................................................................................... 11 E. suncus 12 Blarina E. blarinae 13 E. brevicauda 13 I. brevicauda 14 Cryptotis E.