(Euphorbiaceae) (1807) As Stillingia Populnea. Only By
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

(EUPHORBIACEAE)I [Wood Anatomy of Sapium Haematospermum Müli. Ar
BALDUINIA. n. 35, p. 27-31, 30-V-2012 ESTUDO ANATÔMICO DO LENHO DE SAPIUM HAEMATOSPERMUM MÜLL. ARG (EUPHORBIACEAE)I ANELISE MARTA SIEGLOCH2 JOSÉ NEWTON CARDOSO MARCHIORP SIDINEI RODRIGUES DOS SANTOS4 RESUMO No presente estudo é descrito o lenho de Sapium haematospermum Müll. Arg., com base em material pro- cedente de São Francisco de Assis, Rio Grande do Sul. Foram observadas as seguintes características anatômicas, comuns em Euphorbioideae e gênero Sapium: anéis de crescimento pouco conspícuos; poros de diâmetro médio, pouco numerosos e em curtos múltiplos radiais; placas de perfuração simples; pontoações intervasculares grandes; parênquima apotraqueal difuso-em-agregados; e raios uni e bisseriados, heterocelulares, com cristais e lactíferos. Palavras-chave: Anatomia da madeira, Euphorbioideae, Sapium haemastospermum. ABSTRACT [Wood anatomy of Sapium haematospermum MülI. Arg. (Euphorbiaceae)]. The wood anatomy of Sapium haematospermum Müll. Arg. is described, based on material colleted in the municipality of São Francisco de Assis, Rio Grande do Sul state, Brazil. The following anatomical features that are common among woods of Euphorbioideae and genus Sapium were observed: growth rings almost indistinct; few numerous medium vessels, in short radial multiples; simple perforation plates; large intervascular pits; diffuse-in-aggregates apotracheal parenchyma; and uniseriate and bisseriate heterocellular rays, with crystals and laticifers. Key words: Euphorbioideae, Sapium haematospermum, Wood anatomy. INTRODUÇÃO Gymnanthes, Stillingia e -

Morphology and Anatomy of Foliar Nectaries and Associated Leaves in Mallotus (Euphorbiaceae) Thomas S
Aliso: A Journal of Systematic and Evolutionary Botany Volume 11 | Issue 1 Article 3 1985 Morphology and Anatomy of Foliar Nectaries and Associated Leaves in Mallotus (Euphorbiaceae) Thomas S. Elias Rancho Santa Ana Botanic Garden Sun An-Ci The Chinese Academy of Sciences Follow this and additional works at: http://scholarship.claremont.edu/aliso Part of the Botany Commons Recommended Citation Elias, Thomas S. and An-Ci, Sun (1985) "Morphology and Anatomy of Foliar Nectaries and Associated Leaves in Mallotus (Euphorbiaceae)," Aliso: A Journal of Systematic and Evolutionary Botany: Vol. 11: Iss. 1, Article 3. Available at: http://scholarship.claremont.edu/aliso/vol11/iss1/3 ALISO 11(1),1985, pp. 17-25 MORPHOLOGY AND ANATOMY OF FOLIAR NECTARIES AND ASSOCIATED LEAVES IN MALLOTUS (EUPHORBIACEAE) THOMAS S. ELIAS Rancho Santa Ana Botanic Garden Claremont, California 91711 AND SUN AN-CI Institute of Botany 141 Hsi Chih Men Wai Ta Chie Beijing, People's Republic of China ABSTRACT The morphology and anatomy of the foliar nectaries and associated leaves offour species of Mallotus (Euphorbiaceae) were studied. Light microscopic observations of paraffin- and plastic-embedded spec imens were complemented with scanning electron micrographs. Leaf anatomy of the four species is typical of large mesophytic plants. Aattened foliar nectaries are shown to be composed of specialized epidermal cells. The nonvascularized nectaries consist of narrow columnar cells each with a large nucleus, numerous vacuoles, and dense cytoplasm. Subglandular parenchyma cells have more pro nounced nuclei, more vacuoles and denser cytoplasm than do typical laminar parenchyma. Structurally, these nectaries are similar to those found in other taxa of Euphorbiaceae and in other families of flowering plants. -

Excoecaria Agallocha L
ANNEE : 2015 THESES 2015/ TOU3/ 2020 THESE POUR LE DIPLOME D'ETAT DE DOCTEUR EN PHARMACIE Présentée et soutenue publiquement par ALBINET Lucile Excoecaria agallocha L. Le 16 Mars 2015 Directrice de thèse : VANSTEELANDT Marieke JURY Président : FABRE, Nicolas 1er assesseur : VANSTEELANDT, Marieke 2ème assesseur : AMOUROUX, Noël 1 REMERCIEMENTS Au Président du jury : Mr Fabre, Professeur des Universités à la Faculté des Sciences Pharmaceutiques de Toulouse, je vous remercie de votre disponibilité et d’avoir accepté la présidence de ce jury. Aux membres du jury : Mme Vansteelandt, Maître de Conférences des Universités, et directrice de cette thèse je vous remercie de m’avoir accompagnée dans ce travail et pour votre aide précieuse tout au long de cette thèse. Mr Amouroux, Pharmacien et intervenant à la Faculté de Pharmacie, je vous remercie d’avoir accepté avec spontanéité de faire parti du jury de cette thèse ainsi que pour votre pédagogie tout au long de nos études. A mes parents, qui m’ont toujours encouragée et soutenue. A ma sœur, mon frère, Clau et Jean–Christophe merci pour votre présence et votre soutien. Et comme on dit : « La famille, c’est important la famille ! » A mes amies, Philippine et Sandrine que j’ai eu l’honneur de rencontrer grace à nos études de pharmacie, et dont l’amitié durera encore pour longtemps. Et merci à tous mes amis qui m’accompagnent au quotidien. Enfin, MERCI Rémi pour ton amour et ta grande patience qui, face à cette thèse, ont été mis à rude épreuve mais qui ne t’a pas empêché de vouloir m’épouser ! De tout mon cœur merci. -

Stillingia: a Newly Recorded Genus of Euphorbiaceae from China
Phytotaxa 296 (2): 187–194 ISSN 1179-3155 (print edition) http://www.mapress.com/j/pt/ PHYTOTAXA Copyright © 2017 Magnolia Press Article ISSN 1179-3163 (online edition) https://doi.org/10.11646/phytotaxa.296.2.8 Stillingia: A newly recorded genus of Euphorbiaceae from China SHENGCHUN LI1, 2, BINGHUI CHEN1, XIANGXU HUANG1, XIAOYU CHANG1, TIEYAO TU*1 & DIANXIANG ZHANG1 1 Key Laboratory of Plant Resources Conservation and Sustainable Utilization, South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China 2University of Chinese Academy of Sciences, Beijing 100049, China * Corresponding author, email: [email protected] Abstract Stillingia (Euphorbiaceae) contains ca. 30 species from Latin America, the southern United States, and various islands in the tropical Pacific and in the Indian Ocean. We report here for the first time the occurrence of a member of the genus in China, Stillingia lineata subsp. pacifica. The distribution of the genus in China is apparently narrow, known only from Pingzhou and Wanzhou Islands of the Wanshan Archipelago in the South China Sea, which is close to the Pearl River estuary. This study updates our knowledge on the geographic distribution of the genus, and provides new palynological data as well. Key words: Island, Hippomaneae, South China Sea, Stillingia lineata Introduction During the last decade, hundreds of new plant species or new species records have been added to the flora of China. Nevertheless, newly described or newly recorded plant genera are not discovered and reported very often, suggesting that botanical expedition and plant survey at the generic level may be advanced in China. As far as we know, only six and eight angiosperm genera respectively have been newly described or newly recorded from China within the last ten years (Qiang et al. -

Origin of the Cyathium-Bearing Euphorbieae (Euphorbiaceae): Phylogenetic Study Based on Morphological Characters
ParkBot. Bull.and Backlund Acad. Sin. — (2002) Origin 43: of 57-62 the cyathium-bearing Euphorbieae 57 Origin of the cyathium-bearing Euphorbieae (Euphorbiaceae): phylogenetic study based on morphological characters Ki-Ryong Park1,* and Anders Backlund2 1Department of Biology, Kyung-Nam University, Masan 631-701, Korea 2Division of Pharmacognosy, Department of Pharmacy, Uppsala University, BMC-Biomedical center, S-751 23 Uppsala, Sweden (Received October 6, 2000; Accepted August 24, 2001) Abstract. A cladistic analysis of the subfamily Euphorbioideae was undertaken to elucidate the origin of the cyathium- bearing Euphorbieae and to provide hypotheses about evolutionary relationships within the subfamily. Twenty-one species representing most of the genera within the study group and three outgroup taxa from the subfamilies Acalyphoideae and Crotonoideae were selected for parsimony analysis. An unweighted parsimony analysis of 24 morphological characters resulted in five equally parsimonious trees with consistency indices of 0.67 and tree lengths of 39 steps. The strict consensus tree supported monophyly of the cyathium-bearing Euphorbieae. The sister group relationships of cyathium bearing Euphorbieae with Maprounea (subtribe Hippomaninae) were supported weakly, and the origin of cyathium is possibly in Hippomaneae, or in the common ancestor of Euphorbieae and remaining taxa of Euphorbioideae plus Acalyphoideae. Within the tribe Euphorbieae, both subtribes Euphorbiinae and Neoguilauminiinae are monophyletic, but the African endemic subtribe Anthosteminae is unresolved. The resulting trees support the monophyly of the tribe Stomatocalyceae while the tribe Hippomaneae does not consistently form a clade. Keywords: Cyathium; Euphorbieae; Phylogeny. Introduction to the position of a female flower. Accordingly, the Eu- phorbia-like cyathium results from the alteration of floral In a recent classification of subfamily Euphorbioideae axis and the condensation of the axis of male flower in Boiss., Webster (1975, 1994b) recognized six tribes: Hippomaneae. -

Excoecaria Agallocha L. Antimicrobial Properties Against Important Pathogenic Microorganisms
International Journal of ChemTech Research CODEN( USA): IJCRGG ISSN : 0974-4290 Vol.1, No.4, pp 865-867, Oct-Dec 2009 EXCOECARIA AGALLOCHA L. ANTIMICROBIAL PROPERTIES AGAINST IMPORTANT PATHOGENIC MICROORGANISMS Varahalarao Vadlapudi1, Varaprasad Bobbarala1*, Somasekhar Penumajji2, K. Chandrasekhar Naidu1 1Department of Botany, Andhra University, Visakhapatnam-3, A.P.,India. 2Vivimed labs Limited, 2nd, 4th Floor, Veeranag towers, Habsiguda, Hyderabad, A.P.,India. * Corresponding Author: [email protected] ABSTRACT: Excoecaria agallocha L. leaves were extracted by various extracting procedures, using different solvents for testing the antimicrobial activities against important microorganisms using agar well diffusion method. Chloroform and methanolic extracts were found to be effective against these organisms, whereas hexane extracts were inactive. The purpose of this study was to find preliminary data for the development of alternative treatments to chemical microbicides for the control of plant diseases from natural plant extracts. Keywords: Excoecaria agallocha, Agar well diffusion method; Antimicrobial activity. INTRODUCTION phytochemical and bioactivity studies on mangrove Medical plants have been used for years in daily plants from Kakinada and Godavari, we now report life to treat disease all over the world. It is well known assessment of in vitro antimicrobial activity including that some plants containing active compounds are able to pathogenic bacterial and fungal strains. inhibit the microbial growth. The potential of antimicrobial properties of plants are related to their MATERIALS AND METHODS ability to synthesize compounds by the secondary E. agallocha L. commonly known as milky metabolism. Secondary metabolites proved to be the most mangrove and its vernacular name is Tilla and this important group of compounds that showed wide range species of mangrove tree classified in the plant family of antibacterial and antifungal activity. -

ISB: Atlas of Florida Vascular Plants
Longleaf Pine Preserve Plant List Acanthaceae Asteraceae Wild Petunia Ruellia caroliniensis White Aster Aster sp. Saltbush Baccharis halimifolia Adoxaceae Begger-ticks Bidens mitis Walter's Viburnum Viburnum obovatum Deer Tongue Carphephorus paniculatus Pineland Daisy Chaptalia tomentosa Alismataceae Goldenaster Chrysopsis gossypina Duck Potato Sagittaria latifolia Cow Thistle Cirsium horridulum Tickseed Coreopsis leavenworthii Altingiaceae Elephant's foot Elephantopus elatus Sweetgum Liquidambar styraciflua Oakleaf Fleabane Erigeron foliosus var. foliosus Fleabane Erigeron sp. Amaryllidaceae Prairie Fleabane Erigeron strigosus Simpson's rain lily Zephyranthes simpsonii Fleabane Erigeron vernus Dog Fennel Eupatorium capillifolium Anacardiaceae Dog Fennel Eupatorium compositifolium Winged Sumac Rhus copallinum Dog Fennel Eupatorium spp. Poison Ivy Toxicodendron radicans Slender Flattop Goldenrod Euthamia caroliniana Flat-topped goldenrod Euthamia minor Annonaceae Cudweed Gamochaeta antillana Flag Pawpaw Asimina obovata Sneezeweed Helenium pinnatifidum Dwarf Pawpaw Asimina pygmea Blazing Star Liatris sp. Pawpaw Asimina reticulata Roserush Lygodesmia aphylla Rugel's pawpaw Deeringothamnus rugelii Hempweed Mikania cordifolia White Topped Aster Oclemena reticulata Apiaceae Goldenaster Pityopsis graminifolia Button Rattlesnake Master Eryngium yuccifolium Rosy Camphorweed Pluchea rosea Dollarweed Hydrocotyle sp. Pluchea Pluchea spp. Mock Bishopweed Ptilimnium capillaceum Rabbit Tobacco Pseudognaphalium obtusifolium Blackroot Pterocaulon virgatum -

Tonantzitlolones from Stillingia Lineata Ssp. Lineata As Potential Inhibitors of Chikungunya Virus Sophie Techer, Emmanuelle Girard-Valenciennes, P
Tonantzitlolones from Stillingia lineata ssp. lineata as potential inhibitors of chikungunya virus Sophie Techer, Emmanuelle Girard-Valenciennes, P. Retailleau, J. Neyts, F. Guéritte, P. Leyssen, M. Litaudon, Jacqueline Smadja, Isabelle Grondin To cite this version: Sophie Techer, Emmanuelle Girard-Valenciennes, P. Retailleau, J. Neyts, F. Guéritte, et al.. Tonantz- itlolones from Stillingia lineata ssp. lineata as potential inhibitors of chikungunya virus. Phytochem- istry Letters, Elsevier, 2015, 12, pp.313-319. 10.1016/j.phytol.2015.04.023. hal-01178980 HAL Id: hal-01178980 https://hal.archives-ouvertes.fr/hal-01178980 Submitted on 16 Nov 2016 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Tonantzitlolones from Stillingia lineata ssp. lineata as potential inhibitors of chikungunya virus a a c b c b c S. Techer , E. Girard-Valenciennes , P. Retailleau , J. Neyts , F. Guéritte , P. Leyssen , M. Litaudon , a a, J. Smadja , I. Grondin * a Laboratoire de Chimie des Substances Naturelles et des Sciences des Aliments (LCSNSA), Université de La Réunion, -

Chinese Tallo\Vgets Worse!
The PALMEITO, Fall 1993, Page 3 Chinese Tallo\V Gets Worse! by Greg Jubinsky ",j~ //~ )", \~ ~\ /' . / //I~,-~J," J' ',' ,/ i., ." I . In 1988,an article entitled "Another f,' / i ~ Exotic Nuisance - the Chinese Tal• / ( /',' /' IiI', ~"C"\'" ' ' I low Tree" by Steve Farnsworth, pub• \I (I r;"-\,' ' ' lished in The Palmetto, Winter, 1988• 1 ,i ~ 89, referred to Chinese tallow tree "",",/ (Sapium sebiferum) as the "north Flori• da melaleuca", in reference to "chill• ing" similarities to the south and central Florida pest: its potential for rapid expansion; its promotion by beekeepers as a honey plant; and its ability to spread widely from a single parent tree, especially in moist areas. Chinese tallow tree - Sapium sebiferum (L.)Roxb.- is a member of the Euphorbiaceae, a family of more than 1,000species and varieties, many of which are classified as poisonous. Thegenus Sapium consists of approxi• mately 100 species. Common names for S. sebiferum in the United States include popcorn-tree, Florida aspen, Chinese tallow and Chinese tallow• tree. The latter names arise from the Since the initial introductions, it has yellow to red in the fall. It flowers fact that the outer layer of the seed spread into coastal prairie habitats, and fruits from the time it is about can be used to obtain solid vegetable and is now naturalized in the south• three feet (l m) tall. Inflorescence on tallow. The plant is known as the ern coastal plain from South Carolina some plants are solitary terminating popcorn-tree because of the appear• south to Florida and west to Texas. In branchlets, each narrowly cylindrical, ance of white seeds when the cap• Florida, it is distributed from the two to eight inches (5-20 cm) long. -
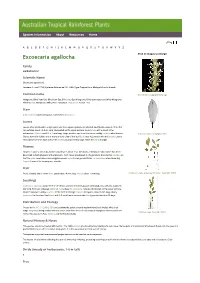
Excoecaria Agallocha Click on Images to Enlarge
Species information Abo ut Reso urces Hom e A B C D E F G H I J K L M N O P Q R S T U V W X Y Z Excoecaria agallocha Click on images to enlarge Family Euphorbiaceae Scientific Name Excoecaria agallocha L. Linnaeus, C. von (1759) Systema Naturae ed. 10 : 1288. Type: Tropical Asia, Malaya & Pacific Islands. Common name Male flowers. Copyright Barry Jago Mangrove, Blind Your Eye; Blind Your Eye; Blind Your Eye Mangrove; Blind-your-eyes-tree; Milky Mangrove; Blinding Tree; Mangrove, Milky; River Poisonous Tree; Scrub Poison Tree Stem Bark exudate rapid and copious. Sometimes deciduous. Leaves Leaves often produced in a tight spiral and thus appear opposite or whorled. Leaf blades about 6-10.5 x 3-5 cm, petioles about 1.5-2 cm long, channelled on the upper surface. Apex retuse with a gland in the indentation. Stipules small, 1.5-2 mm long. Twigs, petioles and leaves produce a milky exudate when broken. Scale bar 10mm. Copyright CSIRO Glands normally visible, one or more on each side of the leaf blade near its junction with the petiole. Lateral veins about 12-18 on each side of the midrib usually forming loops inside the blade margin. Flowers Flowers in spikes, about 40-70 mm long, flowers about 1.5-2 mm diam., emitting an odour which has been described as both pleasant and unpleasant. Each flower enveloped in a bi-glandular bract before anthesis so that the spike resembles a narrow gymnosperm cone. Pollen yellow. Plants +/- deciduous when flowering. -

Fl. China 11: 280–282. 2008. 65. EXCOECARIA Linnaeus, Syst. Nat
Fl. China 11: 280–282. 2008. 65. EXCOECARIA Linnaeus, Syst. Nat., ed. 10, 2: 1288. 1759. 海漆属 hai qi shu Li Bingtao (李秉滔 Li Ping-tao); Hans-Joachim Esser Commia Loureiro. Trees or shrubs, with milky juice, glabrous. Leaves alternate or opposite, petiolate; stipules small, caducous; leaf blade entire or serrulate, penninerved. Flowers unisexual (plants monoecious or dioecious), apetalous, without disk, in axillary or terminal racemelike thyrses. Male flowers (sub)sessile; sepals 3, small, imbricate, free; stamens 3; filaments free; anthers longitudinally dehiscent, without pistillode. Female flowers sessile to pedicellate; calyx 3-lobed or 3-partite; ovary 3-celled, smooth; ovules 1 per locule; stigmas extended or recurved, free to slightly connate at base, undivided, eglandular. Capsules dehiscent into 2-valved cocci; columella persistent, winged. Seeds globose, estrophiolate; episperm crustaceous; endosperm fleshy; cotyledon broad and flattened. About 35 species: Africa, Asia, Australia, Pacific islands; five species (two endemic) in China. 1a. Leaves opposite above, alternate on lower parts. 2a. Leaf blade purple or dark red abaxially........................................................................................................ 1. E. cochinchinensis 2b. Leaf blade green or greenish abaxially when old. 3a. Leaf blade ca. 3 × as long as wide, apex acuminate, not falcate, petiole 3–13 mm ............................ 1. E. cochinchinensis 3b. Leaf blade more than 5 × as long as wide, apex acuminate-falcate, petiole 3–5 mm .................................... 2. E. venenata 1b. Leaves alternate on all parts. 4a. Leaf blade serrulate; male bracts 2- or 3-flowered ............................................................................................... 5. E. acerifolia 4b. Leaf blade entire or nearly so; male bracts 1-flowered. 5a. Petioles 2-glandular at apex; plants dioecious ................................................................................................ 3. E. agallocha 5b. -

The Island Rule and Its Application to Multiple Plant Traits
The island rule and its application to multiple plant traits Annemieke Lona Hedi Hendriks A thesis submitted to the Victoria University of Wellington in partial fulfilment of the requirements for the degree of Master of Science in Ecology and Biodiversity Victoria University of Wellington, New Zealand 2019 ii “The larger the island of knowledge, the longer the shoreline of wonder” Ralph W. Sockman. iii iv General Abstract Aim The Island Rule refers to a continuum of body size changes where large mainland species evolve to become smaller and small species evolve to become larger on islands. Previous work focuses almost solely on animals, with virtually no previous tests of its predictions on plants. I tested for (1) reduced floral size diversity on islands, a logical corollary of the island rule and (2) evidence of the Island Rule in plant stature, leaf size and petiole length. Location Small islands surrounding New Zealand; Antipodes, Auckland, Bounty, Campbell, Chatham, Kermadec, Lord Howe, Macquarie, Norfolk, Snares, Stewart and the Three Kings. Methods I compared the morphology of 65 island endemics and their closest ‘mainland’ relative. Species pairs were identified. Differences between archipelagos located at various latitudes were also assessed. Results Floral sizes were reduced on islands relative to the ‘mainland’, consistent with predictions of the Island Rule. Plant stature, leaf size and petiole length conformed to the Island Rule, with smaller plants increasing in size, and larger plants decreasing in size. Main conclusions Results indicate that the conceptual umbrella of the Island Rule can be expanded to plants, accelerating understanding of how plant traits evolve on isolated islands.