Oceanographic Considerations for Exoplanet Life Detection
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Fronts in the World Ocean's Large Marine Ecosystems. ICES CM 2007
- 1 - This paper can be freely cited without prior reference to the authors International Council ICES CM 2007/D:21 for the Exploration Theme Session D: Comparative Marine Ecosystem of the Sea (ICES) Structure and Function: Descriptors and Characteristics Fronts in the World Ocean’s Large Marine Ecosystems Igor M. Belkin and Peter C. Cornillon Abstract. Oceanic fronts shape marine ecosystems; therefore front mapping and characterization is one of the most important aspects of physical oceanography. Here we report on the first effort to map and describe all major fronts in the World Ocean’s Large Marine Ecosystems (LMEs). Apart from a geographical review, these fronts are classified according to their origin and physical mechanisms that maintain them. This first-ever zero-order pattern of the LME fronts is based on a unique global frontal data base assembled at the University of Rhode Island. Thermal fronts were automatically derived from 12 years (1985-1996) of twice-daily satellite 9-km resolution global AVHRR SST fields with the Cayula-Cornillon front detection algorithm. These frontal maps serve as guidance in using hydrographic data to explore subsurface thermohaline fronts, whose surface thermal signatures have been mapped from space. Our most recent study of chlorophyll fronts in the Northwest Atlantic from high-resolution 1-km data (Belkin and O’Reilly, 2007) revealed a close spatial association between chlorophyll fronts and SST fronts, suggesting causative links between these two types of fronts. Keywords: Fronts; Large Marine Ecosystems; World Ocean; sea surface temperature. Igor M. Belkin: Graduate School of Oceanography, University of Rhode Island, 215 South Ferry Road, Narragansett, Rhode Island 02882, USA [tel.: +1 401 874 6533, fax: +1 874 6728, email: [email protected]]. -

A Destabilizing Thermohaline Circulation–Atmosphere–Sea Ice
642 JOURNAL OF CLIMATE VOLUME 12 NOTES AND CORRESPONDENCE A Destabilizing Thermohaline Circulation±Atmosphere±Sea Ice Feedback STEVEN R. JAYNE MIT±WHOI Joint Program in Oceanography, Woods Hole Oceanographic Institution, Woods Hole, Massachusetts JOCHEM MAROTZKE Center for Global Change Science, Department of Earth, Atmospheric and Planetary Sciences, Massachusetts Institute of Technology, Cambridge, Massachusetts 18 November 1996 and 9 March 1998 ABSTRACT Some of the interactions and feedbacks between the atmosphere, thermohaline circulation, and sea ice are illustrated using a simple process model. A simpli®ed version of the annual-mean coupled ocean±atmosphere box model of Nakamura, Stone, and Marotzke is modi®ed to include a parameterization of sea ice. The model includes the thermodynamic effects of sea ice and allows for variable coverage. It is found that the addition of sea ice introduces feedbacks that have a destabilizing in¯uence on the thermohaline circulation: Sea ice insulates the ocean from the atmosphere, creating colder air temperatures at high latitudes, which cause larger atmospheric eddy heat and moisture transports and weaker oceanic heat transports. These in turn lead to thicker ice coverage and hence establish a positive feedback. The results indicate that generally in colder climates, the presence of sea ice may lead to a signi®cant destabilization of the thermohaline circulation. Brine rejection by sea ice plays no important role in this model's dynamics. The net destabilizing effect of sea ice in this model is the result of two positive feedbacks and one negative feedback and is shown to be model dependent. To date, the destabilizing feedback between atmospheric and oceanic heat ¯uxes, mediated by sea ice, has largely been neglected in conceptual studies of thermohaline circulation stability, but it warrants further investigation in more realistic models. -

World Ocean Thermocline Weakening and Isothermal Layer Warming
applied sciences Article World Ocean Thermocline Weakening and Isothermal Layer Warming Peter C. Chu * and Chenwu Fan Naval Ocean Analysis and Prediction Laboratory, Department of Oceanography, Naval Postgraduate School, Monterey, CA 93943, USA; [email protected] * Correspondence: [email protected]; Tel.: +1-831-656-3688 Received: 30 September 2020; Accepted: 13 November 2020; Published: 19 November 2020 Abstract: This paper identifies world thermocline weakening and provides an improved estimate of upper ocean warming through replacement of the upper layer with the fixed depth range by the isothermal layer, because the upper ocean isothermal layer (as a whole) exchanges heat with the atmosphere and the deep layer. Thermocline gradient, heat flux across the air–ocean interface, and horizontal heat advection determine the heat stored in the isothermal layer. Among the three processes, the effect of the thermocline gradient clearly shows up when we use the isothermal layer heat content, but it is otherwise when we use the heat content with the fixed depth ranges such as 0–300 m, 0–400 m, 0–700 m, 0–750 m, and 0–2000 m. A strong thermocline gradient exhibits the downward heat transfer from the isothermal layer (non-polar regions), makes the isothermal layer thin, and causes less heat to be stored in it. On the other hand, a weak thermocline gradient makes the isothermal layer thick, and causes more heat to be stored in it. In addition, the uncertainty in estimating upper ocean heat content and warming trends using uncertain fixed depth ranges (0–300 m, 0–400 m, 0–700 m, 0–750 m, or 0–2000 m) will be eliminated by using the isothermal layer. -

Downloaded from the Coriolis Global Data Center in France (Ftp://Ftp.Ifremer.Fr)
remote sensing Article Impact of Enhanced Wave-Induced Mixing on the Ocean Upper Mixed Layer during Typhoon Nepartak in a Regional Model of the Northwest Pacific Ocean Chengcheng Yu 1 , Yongzeng Yang 2,3,4, Xunqiang Yin 2,3,4,*, Meng Sun 2,3,4 and Yongfang Shi 2,3,4 1 Ocean College, Zhejiang University, Zhoushan 316000, China; [email protected] 2 First Institute of Oceanography, Ministry of Natural Resources, Qingdao 266061, China; yangyz@fio.org.cn (Y.Y.); sunm@fio.org.cn (M.S.); shiyf@fio.org.cn (Y.S.) 3 Laboratory for Regional Oceanography and Numerical Modeling, Pilot National Laboratory for Marine Science and Technology, Qingdao 266071, China 4 Key Laboratory of Marine Science and Numerical Modeling (MASNUM), Ministry of Natural Resources, Qingdao 266061, China * Correspondence: yinxq@fio.org.cn Received: 30 July 2020; Accepted: 27 August 2020; Published: 30 August 2020 Abstract: To investigate the effect of wave-induced mixing on the upper ocean structure, especially under typhoon conditions, an ocean-wave coupled model is used in this study. Two physical processes, wave-induced turbulence mixing and wave transport flux residue, are introduced. We select tropical cyclone (TC) Nepartak in the Northwest Pacific ocean as a TC example. The results show that during the TC period, the wave-induced turbulence mixing effectively increases the cooling area and cooling amplitude of the sea surface temperature (SST). The wave transport flux residue plays a positive role in reproducing the distribution of the SST cooling area. From the intercomparisons among experiments, it is also found that the wave-induced turbulence mixing has an important effect on the formation of mixed layer depth (MLD). -

Influences of the Ocean Surface Mixed Layer and Thermohaline Stratification on Arctic Sea Ice in the Central Canada Basin J
JOURNAL OF GEOPHYSICAL RESEARCH, VOL. 115, C10018, doi:10.1029/2009JC005660, 2010 Influences of the ocean surface mixed layer and thermohaline stratification on Arctic Sea ice in the central Canada Basin J. M. Toole,1 M.‐L. Timmermans,2 D. K. Perovich,3 R. A. Krishfield,1 A. Proshutinsky,1 and J. A. Richter‐Menge3 Received 22 July 2009; revised 19 April 2010; accepted 1 June 2010; published 8 October 2010. [1] Variations in the Arctic central Canada Basin mixed layer properties are documented based on a subset of nearly 6500 temperature and salinity profiles acquired by Ice‐ Tethered Profilers during the period summer 2004 to summer 2009 and analyzed in conjunction with sea ice observations from ice mass balance buoys and atmosphere‐ocean heat flux estimates. The July–August mean mixed layer depth based on the Ice‐Tethered Profiler data averaged 16 m (an overestimate due to the Ice‐Tethered Profiler sampling characteristics and present analysis procedures), while the average winter mixed layer depth was only 24 m, with individual observations rarely exceeding 40 m. Guidance interpreting the observations is provided by a 1‐D ocean mixed layer model. The analysis focuses attention on the very strong density stratification at the base of the mixed layer in the Canada Basin that greatly impedes surface layer deepening and thus limits the flux of deep ocean heat to the surface that could influence sea ice growth/ decay. The observations additionally suggest that efficient lateral mixed layer restratification processes are active in the Arctic, also impeding mixed layer deepening. Citation: Toole, J. -

Glacial Ocean Circulation and Stratification Explained by Reduced
Glacial ocean circulation and stratification explained by reduced atmospheric temperature Malte F. Jansena,1 aDepartment of the Geophysical Sciences, The University of Chicago, Chicago, IL 60637 Edited by Mark H. Thiemens, University of California, San Diego, La Jolla, CA, and approved November 7, 2016 (received for review June 27, 2016) Earth’s climate has undergone dramatic shifts between glacial and We test the connection between atmospheric temperature interglacial time periods, with high-latitude temperature changes and ocean circulation and stratification changes, using idealized on the order of 5–10 ◦C. These climatic shifts have been asso- numerical simulations, which allow us to isolate the proposed ciated with major rearrangements in the deep ocean circulation mechanism. We use a coupled ocean–sea-ice model, with atmo- and stratification, which have likely played an important role in spheric temperature, winds, and evaporation–precipitation pre- the observed atmospheric carbon dioxide swings by affecting the scribed as boundary conditions (Materials and Methods). The partitioning of carbon between the atmosphere and the ocean. model uses an idealized continental configuration resembling the The mechanisms by which the deep ocean circulation changed, Atlantic and Southern Oceans, where the most elemental circu- however, are still unclear and represent a major challenge to our lation changes have been inferred (3, 5, 6, 12). understanding of glacial climates. This study shows that vari- ous inferred changes in the deep ocean circulation and stratifica- Results tion between glacial and interglacial climates can be interpreted We first focus on the model’s ability to reproduce key features as a direct consequence of atmospheric temperature differences. -

Ice Production in Ross Ice Shelf Polynyas During 2017–2018 from Sentinel–1 SAR Images
remote sensing Article Ice Production in Ross Ice Shelf Polynyas during 2017–2018 from Sentinel–1 SAR Images Liyun Dai 1,2, Hongjie Xie 2,3,* , Stephen F. Ackley 2,3 and Alberto M. Mestas-Nuñez 2,3 1 Key Laboratory of Remote Sensing of Gansu Province, Heihe Remote Sensing Experimental Research Station, Cold and Arid Regions Environmental and Engineering Research Institute, Chinese Academy of Sciences, Lanzhou 730000, China; [email protected] 2 Laboratory for Remote Sensing and Geoinformatics, Department of Geological Sciences, University of Texas at San Antonio, San Antonio, TX 78249, USA; [email protected] (S.F.A.); [email protected] (A.M.M.-N.) 3 Center for Advanced Measurements in Extreme Environments, University of Texas at San Antonio, San Antonio, TX 78249, USA * Correspondence: [email protected]; Tel.: +1-210-4585445 Received: 21 April 2020; Accepted: 5 May 2020; Published: 7 May 2020 Abstract: High sea ice production (SIP) generates high-salinity water, thus, influencing the global thermohaline circulation. Estimation from passive microwave data and heat flux models have indicated that the Ross Ice Shelf polynya (RISP) may be the highest SIP region in the Southern Oceans. However, the coarse spatial resolution of passive microwave data limited the accuracy of these estimates. The Sentinel-1 Synthetic Aperture Radar dataset with high spatial and temporal resolution provides an unprecedented opportunity to more accurately distinguish both polynya area/extent and occurrence. In this study, the SIPs of RISP and McMurdo Sound polynya (MSP) from 1 March–30 November 2017 and 2018 are calculated based on Sentinel-1 SAR data (for area/extent) and AMSR2 data (for ice thickness). -

Antarctic Sea Ice Control on Ocean Circulation in Present and Glacial Climates
Antarctic sea ice control on ocean circulation in present and glacial climates Raffaele Ferraria,1, Malte F. Jansenb, Jess F. Adkinsc, Andrea Burkec, Andrew L. Stewartc, and Andrew F. Thompsonc aDepartment of Earth, Atmospheric and Planetary Sciences, Massachusetts Institute of Technology, Cambridge, MA 02139; bAtmospheric and Oceanic Sciences Program, Geophysical Fluid Dynamics Laboratory, Princeton, NJ 08544; and cDivision of Geological and Planetary Sciences, California Institute of Technology, Pasadena, CA 91125 Edited* by Edward A. Boyle, Massachusetts Institute of Technology, Cambridge, MA, and approved April 16, 2014 (received for review December 31, 2013) In the modern climate, the ocean below 2 km is mainly filled by waters possibly associated with an equatorward shift of the Southern sinking into the abyss around Antarctica and in the North Atlantic. Hemisphere westerlies (11–13), (ii) an increase in abyssal stratifi- Paleoproxies indicate that waters of North Atlantic origin were instead cation acting as a lid to deep carbon (14), (iii)anexpansionofseaice absent below 2 km at the Last Glacial Maximum, resulting in an that reduced the CO2 outgassing over the Southern Ocean (15), and expansion of the volume occupied by Antarctic origin waters. In this (iv) a reduction in the mixing between waters of Antarctic and Arctic study we show that this rearrangement of deep water masses is origin, which is a major leak of abyssal carbon in the modern climate dynamically linked to the expansion of summer sea ice around (16). Current understanding is that some combination of all of these Antarctica. A simple theory further suggests that these deep waters feedbacks, together with a reorganization of the biological and only came to the surface under sea ice, which insulated them from carbonate pumps, is required to explain the observed glacial drop in atmospheric forcing, and were weakly mixed with overlying waters, atmospheric CO2 (17). -

Coastal Upwelling Revisited: Ekman, Bakun, and Improved 10.1029/2018JC014187 Upwelling Indices for the U.S
Journal of Geophysical Research: Oceans RESEARCH ARTICLE Coastal Upwelling Revisited: Ekman, Bakun, and Improved 10.1029/2018JC014187 Upwelling Indices for the U.S. West Coast Key Points: Michael G. Jacox1,2 , Christopher A. Edwards3 , Elliott L. Hazen1 , and Steven J. Bograd1 • New upwelling indices are presented – for the U.S. West Coast (31 47°N) to 1NOAA Southwest Fisheries Science Center, Monterey, CA, USA, 2NOAA Earth System Research Laboratory, Boulder, CO, address shortcomings in historical 3 indices USA, University of California, Santa Cruz, CA, USA • The Coastal Upwelling Transport Index (CUTI) estimates vertical volume transport (i.e., Abstract Coastal upwelling is responsible for thriving marine ecosystems and fisheries that are upwelling/downwelling) disproportionately productive relative to their surface area, particularly in the world’s major eastern • The Biologically Effective Upwelling ’ Transport Index (BEUTI) estimates boundary upwelling systems. Along oceanic eastern boundaries, equatorward wind stress and the Earth s vertical nitrate flux rotation combine to drive a near-surface layer of water offshore, a process called Ekman transport. Similarly, positive wind stress curl drives divergence in the surface Ekman layer and consequently upwelling from Supporting Information: below, a process known as Ekman suction. In both cases, displaced water is replaced by upwelling of relatively • Supporting Information S1 nutrient-rich water from below, which stimulates the growth of microscopic phytoplankton that form the base of the marine food web. Ekman theory is foundational and underlies the calculation of upwelling indices Correspondence to: such as the “Bakun Index” that are ubiquitous in eastern boundary upwelling system studies. While generally M. G. Jacox, fi [email protected] valuable rst-order descriptions, these indices and their underlying theory provide an incomplete picture of coastal upwelling. -
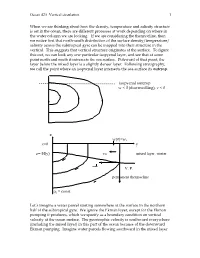
Ocean 423 Vertical Circulation 1 When We Are Thinking About How The
Ocean 423 Vertical circulation 1 When we are thinking about how the density, temperature and salinity structure is set in the ocean, there are different processes at work depending on where in the water column we are looking. If we are considering the thermocline, then we notice first that north-south distribution of the surface density/temperature/ salinity across the subtropical gyre can be mapped into their structure in the vertical. This suggests that vertical structure originates at the surface. To figure this out, we can look any one particular isopycnal layer, and see that at some point north and south it intersects the sea surface. Poleward of that point, the layer below the mixed layer is a slightly denser layer. Following stratigraphy, we call the point where an isopycnal layer intersects the sea-surface its outcrop. isopycnal outcrop w < 0 (downwelling), v < 0 z w(0)=we z=0 y z=-H(y) vG mixed layer, winter L.I. V. P. permanent thermocline = const Let’s imagine a water parcel starting somewhere at the surface in the northern half of the subtropical gyre. We ignore the Ekman layer, except for the Ekman pumping it produces, which we specify as a boundary condition on vertical velocity at the ocean surface. The geostrophic velocity is southward everywhere (including the mixed layer) in this part of the ocean because of the downward Ekman pumping. Imagine water parcels flowing southward in the mixed layer Ocean 423 Vertical circulation 2 (and no heating or cooling). A water parcel, when it encounters lighter water to the south will tend to follow its own isopycnal down out of the mixed layer. -

Implementation of an Ocean Mixed Layer Model in IFS
622 Implementation of an ocean mixed layer model in IFS Y. Takaya12, F. Vitart1, G. Balsamo1, M. Balmaseda1, M. Leutbecher1, F. Molteni1 Research Department 1European Centre for Medium-Range Weather Forecasts, Reading, UK 2Japan Meteorological Agency, Tokyo, Japan March 2010 Series: ECMWF Technical Memoranda A full list of ECMWF Publications can be found on our web site under: http://www.ecmwf.int/publications/ Contact: [email protected] c Copyright 2010 European Centre for Medium-Range Weather Forecasts Shinfield Park, Reading, RG2 9AX, England Literary and scientific copyrights belong to ECMWF and are reserved in all countries. This publication is not to be reprinted or translated in whole or in part without the written permission of the Director. Appropriate non-commercial use will normally be granted under the condition that reference is made to ECMWF. The information within this publication is given in good faith and considered to be true, but ECMWF accepts no liability for error, omission and for loss or damage arising from its use. Implementation of an ocean mixed layer model Abstract An ocean mixed layer model has been implemented in the ECMWF Integrated Forecasting System (IFS) in order to have a better representation of the atmosphere-upper ocean coupled processes in the ECMWF medium-range forecasts. The ocean mixed layer model uses a non-local K profile parameterization (KPP; Large et al. 1994). This model is a one-dimensional column model with high vertical resolution near the surface to simulate the diurnal thermal variations in the upper ocean. Its simplified dynamics makes it cheaper than a full ocean general circulation model and its implementation in IFS allows the atmosphere- ocean coupled model to run much more efficiently than using a coupler. -

DESALINATION: Balancing the Socioeconomic Benefits and Environmental Costs
DESALINATION: Balancing the Socioeconomic Benefits and Environmental Costs www.research.natixis.com https://gsh.cib.natixis.com executive summary Chapter 1 Making sense of desalination: technological, financial and economic aspects of desalination assets Chapter 2 Sustainability assessment of desalination assets: recognizing the socioeconomic benefits and mitigating environmental costs of desalination Chapter 3 Desalination sustainability performance scorecard acknowledgements appendix biblioghraphy TABLE OF CONTENTS OF TABLE 1. Making sense of desalination: technological, financial and economic aspects of desalination assets 1. DESALINATION TECHNOLOGIES 2. FINANCIAL AND ECONOMIC ASPECTS OF DESALINATION ASSETS 1.1. AN OVERVIEW OF DESALINATION TECHNOLOGIES 2.1.THE DEVELOPMENT AND FINANCING OF DESALINATION ASSETS Thermal desalination: Multistage Flash Distillation and Multieffect Distillation Building and operating desalination assets: complex and evolving value chain Membrane desalination: Reverse Osmosis Project development models: fine-tuning Hybridization of thermal and the appropriate risk-sharing model membrane desalination Bringing capital to desalination assets: A set of parameters to assess the performance an increasingly strategic issue and efficiency of desalination assets Case study of desalination in Israel: innovative 1.2. A BRIEF HISTORY AND financing schemes achieving some of the GEOGRAPHICAL DISTRIBUTION OF lowest desalinated water costs worldwide DESALINATION TECHNOLOGIES Case study of desalination in Singapore: The market