Differential Mechanisms of Action of the Trace Amines Octopamine
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
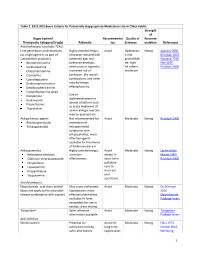
Table 2. 2012 AGS Beers Criteria for Potentially
Table 2. 2012 AGS Beers Criteria for Potentially Inappropriate Medication Use in Older Adults Strength of Organ System/ Recommendat Quality of Recomm Therapeutic Category/Drug(s) Rationale ion Evidence endation References Anticholinergics (excludes TCAs) First-generation antihistamines Highly anticholinergic; Avoid Hydroxyzin Strong Agostini 2001 (as single agent or as part of clearance reduced with e and Boustani 2007 combination products) advanced age, and promethazi Guaiana 2010 Brompheniramine tolerance develops ne: high; Han 2001 Carbinoxamine when used as hypnotic; All others: Rudolph 2008 Chlorpheniramine increased risk of moderate Clemastine confusion, dry mouth, Cyproheptadine constipation, and other Dexbrompheniramine anticholinergic Dexchlorpheniramine effects/toxicity. Diphenhydramine (oral) Doxylamine Use of diphenhydramine in Hydroxyzine special situations such Promethazine as acute treatment of Triprolidine severe allergic reaction may be appropriate. Antiparkinson agents Not recommended for Avoid Moderate Strong Rudolph 2008 Benztropine (oral) prevention of Trihexyphenidyl extrapyramidal symptoms with antipsychotics; more effective agents available for treatment of Parkinson disease. Antispasmodics Highly anticholinergic, Avoid Moderate Strong Lechevallier- Belladonna alkaloids uncertain except in Michel 2005 Clidinium-chlordiazepoxide effectiveness. short-term Rudolph 2008 Dicyclomine palliative Hyoscyamine care to Propantheline decrease Scopolamine oral secretions. Antithrombotics Dipyridamole, oral short-acting* May -

Neurotransmitter Resource Guide
NEUROTRANSMITTER RESOURCE GUIDE Science + Insight doctorsdata.com Doctor’s Data, Inc. Neurotransmitter RESOURCE GUIDE Table of Contents Sample Report Sample Report ........................................................................................................................................................................... 1 Analyte Considerations Phenylethylamine (B-phenylethylamine or PEA) ................................................................................................. 1 Tyrosine .......................................................................................................................................................................................... 3 Tyramine ........................................................................................................................................................................................4 Dopamine .....................................................................................................................................................................................6 3, 4-Dihydroxyphenylacetic Acid (DOPAC) ............................................................................................................... 7 3-Methoxytyramine (3-MT) ............................................................................................................................................... 9 Norepinephrine ........................................................................................................................................................................ -

Methylphenidate Hydrochloride
Application for Inclusion to the 22nd Expert Committee on the Selection and Use of Essential Medicines: METHYLPHENIDATE HYDROCHLORIDE December 7, 2018 Submitted by: Patricia Moscibrodzki, M.P.H., and Craig L. Katz, M.D. The Icahn School of Medicine at Mount Sinai Graduate Program in Public Health New York NY, United States Contact: [email protected] TABLE OF CONTENTS Page 3 Summary Statement Page 4 Focal Point Person in WHO Page 5 Name of Organizations Consulted Page 6 International Nonproprietary Name Page 7 Formulations Proposed for Inclusion Page 8 International Availability Page 10 Listing Requested Page 11 Public Health Relevance Page 13 Treatment Details Page 19 Comparative Effectiveness Page 29 Comparative Safety Page 41 Comparative Cost and Cost-Effectiveness Page 45 Regulatory Status Page 48 Pharmacoepial Standards Page 49 Text for the WHO Model Formulary Page 52 References Page 61 Appendix – Letters of Support 2 1. Summary Statement of the Proposal for Inclusion of Methylphenidate Methylphenidate (MPH), a central nervous system (CNS) stimulant, of the phenethylamine class, is proposed for inclusion in the WHO Model List of Essential Medications (EML) & the Model List of Essential Medications for Children (EMLc) for treatment of Attention-Deficit/Hyperactivity Disorder (ADHD) under ICD-11, 6C9Z mental, behavioral or neurodevelopmental disorder, disruptive behavior or dissocial disorders. To date, the list of essential medications does not include stimulants, which play a critical role in the treatment of psychotic disorders. Methylphenidate is proposed for inclusion on the complimentary list for both children and adults. This application provides a systematic review of the use, efficacy, safety, availability, and cost-effectiveness of methylphenidate compared with other stimulant (first-line) and non-stimulant (second-line) medications. -

Review Paper Monoamine Oxidase Inhibitors: a Review Concerning Dietary Tyramine and Drug Interactions
PsychoTropical Commentaries (2016) 1:1 – 90 © Fernwell Publications Review Paper Monoamine Oxidase Inhibitors: a Review Concerning Dietary Tyramine and Drug Interactions PK Gillman PsychoTropical Research, Bucasia, Queensland, Australia Abstract This comprehensive monograph surveys original data on the subject of both dietary tyramine and drug interactions relevant to Monoamine Oxidase Inhibitors (MAOIs), about which there is much outdated, incorrect and incomplete information in the medical literature and elsewhere. Fewer foods than previously supposed have problematically high tyramine levels because international food hygiene regulations have improved both production and handling. Cheese is the only food that has, in the past, been associated with documented fatalities from hypertension, and now almost all ‘supermarket’ cheeses are perfectly safe in healthy-sized portions. The variability of sensitivity to tyramine between individuals, and the sometimes unpredictable amount of tyramine content in foods, means a little knowledge and care are still advised. The interactions between MAOIs and other drugs are now well understood, are quite straightforward, and are briefly summarized here (by a recognised expert). MAOIs have no apparently clinically relevant pharmaco-kinetic interactions, and the only significant pharmaco-dynamic interaction, other than the ‘cheese reaction’ (caused by indirect sympatho-mimetic activity [ISA], is serotonin toxicity ST (aka serotonin syndrome) which is now well defined and straightforward to avoid by not co-administering any drug with serotonin re-uptake inhibitor (SRI) potency. There are no therapeutically used drugs, other than SRIs, that are capable of inducing serious ST with MAOIs. Anaesthesia is not contra- indicated if a patient is taking MAOIs. Most of the previously held concerns about MAOIs turn out to be mythical: they are either incorrect, or over-rated in importance, or stem from apprehensions born out of insufficient knowledge. -

Download Product Insert (PDF)
PRODUCT INFORMATION Tyramine Item No. 18601 CAS Registry No.: 51-67-2 Formal Name: 4-(2-aminoethyl)-phenol Synonyms: 2-(4-Hydroxyphenyl)ethylamine, NSC 249188, p-Tyramine, NH2 Uteramine MF: C8H11NO FW: 137.2 HO Purity: ≥98% UV/Vis.: λmax: 224, 278 nm Supplied as: A crystalline solid Storage: -20°C Stability: ≥2 years Information represents the product specifications. Batch specific analytical results are provided on each certificate of analysis. Laboratory Procedures Tyramine is supplied as a crystalline solid. A stock solution may be made by dissolving the tyramine in the solvent of choice. Tyramine is soluble in organic solvents such as ethanol, DMSO, and dimethyl formamide (DMF), which should be purged with an inert gas. The solubility of tyramine in these solvents is approximately 5, 20, and 25 mg/ml, respectively. Tyramine is sparingly soluble in aqueous buffers. For maximum solubility in aqueous buffers, tyramine should first be dissolved in DMF and then diluted with the aqueous buffer of choice. Tyramine has a solubility of approximately 0.5 mg/ml in a 1:1 solution of DMF:PBS (pH 7.2) using this method. We do not recommend storing the aqueous solution for more than one day. Description Tyramine is a tyrosine-derived endogenous and dietary monoamine and trace amine-associated receptor 1-3 1 (TAAR1) agonist. It activates TAAR1 (EC50s = 0.08, 0.69, and 2.26 µM for rat, mouse, and human-rat chimera receptors, respectively).1 Tyramine also inhibits the release of norepinephrine and dopamine in 4 isolated rat caudate nucleus (IC50s = 40.6 and 119 nM, respectively). -

Appendix A: Potentially Inappropriate Prescriptions (Pips) for Older People (Modified from ‘STOPP/START 2’ O’Mahony Et Al 2014)
Appendix A: Potentially Inappropriate Prescriptions (PIPs) for older people (modified from ‘STOPP/START 2’ O’Mahony et al 2014) Consider holding (or deprescribing - consult with patient): 1. Any drug prescribed without an evidence-based clinical indication 2. Any drug prescribed beyond the recommended duration, where well-defined 3. Any duplicate drug class (optimise monotherapy) Avoid hazardous combinations e.g.: 1. The Triple Whammy: NSAID + ACE/ARB + diuretic in all ≥ 65 year olds (NHS Scotland 2015) 2. Sick Day Rules drugs: Metformin or ACEi/ARB or a diuretic or NSAID in ≥ 65 year olds presenting with dehydration and/or acute kidney injury (AKI) (NHS Scotland 2015) 3. Anticholinergic Burden (ACB): Any additional medicine with anticholinergic properties when already on an Anticholinergic/antimuscarinic (listed overleaf) in > 65 year olds (risk of falls, increased anticholinergic toxicity: confusion, agitation, acute glaucoma, urinary retention, constipation). The following are known to contribute to the ACB: Amantadine Antidepressants, tricyclic: Amitriptyline, Clomipramine, Dosulepin, Doxepin, Imipramine, Nortriptyline, Trimipramine and SSRIs: Fluoxetine, Paroxetine Antihistamines, first generation (sedating): Clemastine, Chlorphenamine, Cyproheptadine, Diphenhydramine/-hydrinate, Hydroxyzine, Promethazine; also Cetirizine, Loratidine Antipsychotics: especially Clozapine, Fluphenazine, Haloperidol, Olanzepine, and phenothiazines e.g. Prochlorperazine, Trifluoperazine Baclofen Carbamazepine Disopyramide Loperamide Oxcarbazepine Pethidine -

TR-348: Alpha-Methyldopa Sesquihydrate (CASRN 41372-08-1)
NATIONAL TOXICOLOGY PROGRAM Technical Report Series No. 348 TOXICOLOGY AND CARCINOGENESIS STUDIES OF a/pha-METHYLDOPA SESQUIHYDRATE (CAS NO. 41372-08-1) IN F344/N RATS AND B6C3Fi MICE (FEED STUDIES) U.S. DEPARTMENT OF HEALTH AND HUMAN SERVICES Public Health Service National Institutes of Health NTP TECHNICAL REPORT ON THE TOXICOLOGY AND CARCINOGENESIS STUDIES OF a/p/)a-METHYLDOPA SESQUIHYDRATE (CAS NO. 41372-08-1) IN F344/N RATS AND B6C3Fi MICE (FEED STUDIES) June K. Dunnick, Ph.D., Chemical Manager NATIONAL TOXICOLOGY PROGRAM P.O. Box 12233 Research Triangle Park, NC 27709 March 1989 NTP TR 348 NIH Publication No. 89-2803 U.S. DEPARTMENT OF HEALTH AND HUMAN SERVICES Public Health Service National Institutes of Health NOTE TO THE READER This study was performed under the direction of the K’ational Institute of Environmental Health Sci- ences as a function of the National Toxicology Program. The studies described in this Technical Re- port have been conducted in compliance with NTP chemical health and safety requirements and must meet or exceed all applicable Federal, state, and local health and safety regulations. Animal care and use were in accordance with the U.S. Public Health Service Policy on Humane Care and Use of Ani- mals. All NTP toxicology and carcinogenesis studies are subjected to a data audit before being pre- sented for public peer review. Although every effort is made to prepare the Technical Reports as accurately as possible, mistakes may occur. Readers are requested to identify any mistakes so that corrective action may be taken. Further, anyone who is aware of related ongoing or published studies not mentioned in this report is encouraged to make this information known to the NTP. -

DART-MS/MS Screening for the Determination of 1,3- Dimethylamylamine and Undeclared Stimulants in Seized Dietary Supplements from Brazil
Forensic Chemistry 8 (2018) 134–145 Contents lists available at ScienceDirect Forensic Chemistry journal homepage: www.elsevier.com/locate/forc DART-MS/MS screening for the determination of 1,3- dimethylamylamine and undeclared stimulants in seized dietary supplements from Brazil Maíra Kerpel dos Santos a, Emily Gleco b, J. Tyler Davidson b, Glen P. Jackson b, Renata Pereira Limberger a, ⇑ Luis E. Arroyo b, a Graduate Program of Pharmaceutical Sciences, Faculty of Pharmacy, Federal University of Rio Grande do Sul, Brazil b Department of Forensic & Investigative Science, West Virginia University, USA article info abstract Article history: 1,3-dimethylamylamine (DMAA) is an alkylamine with stimulating properties that has been used pre- Received 15 December 2017 dominantly as an additive in dietary supplements. DMAA is mostly consumed by professional athletes, Received in revised form 14 March 2018 and several doping cases reported since 2008 led to its prohibition by the World Anti-Doping Agency Accepted 18 March 2018 (WADA) in 2010. Adverse effects have indicated DMAA toxicity, and there is few data regarding its safety, Available online 22 March 2018 so it was banned by regulatory agencies from Brazil and the United States. Ambient ionization methods such as Direct Analysis in Real Time Tandem Mass Spectrometry (DART-MS/MS) are an alternative for Keywords: dietary supplements analysis, because they enable the analysis of samples at atmospheric pressure in 1,3-dimethylamylamine a very short time and with only minimal sample preparation. Therefore, the aim of this work was to DART-MS/MS Dietary supplements develop a methodology by DART-MS/MS to detect the presence of DMAA, ephedrine, synephrine, caffeine, Stimulants sibutramine, and methylphenidate in 108 dietary supplements seized by the Brazilian Federal Police Adulterants (BFP). -

Is TAAR1 a Potential Therapeutic Target for Immune Dysregulation In
Graduate Physical and Life Sciences PhD Pharmacology Abstract ID# 1081 Is TAAR1 a Potential Therapeutic Target for Immune Dysregulation in Drug Abuse? Fleischer, Lisa M; Tamashunas, Nina and Miller, Gregory M Addiction Sciences Laboratory, Northeastern University, Boston MA 02115 Abstract Discovered in 2001, Trace Amine Associated Receptor 1 (TAAR1) is a direct target of Data and Results amphetamine, methamphetamine and MDMA. It is expressed in the brain reward circuity and modulates dopamine transporter function and dopamine neuron firing rates. Newly-developed compounds that specifically target TAAR1 have recently been investigated in animal models In addition to brain, TAAR1 is expressed in immune cells METH promotes PKA and PKC Phosphorylation through TAAR1 as candidate therapeutics for methamphetamine, cocaine and alcohol abuse. These studies • We treated HEK/TAAR1 cells and HEK293 involving classic behavioral measures of drug response, as well as drug self-administration, Rhesus and Human cells with vehicle or METH, with and without strongly implicate TAAR1 as a potential therapeutic target for the treatment of addiction. In activators and inhibitors of PKA and PKC. addition to its central actions, we demonstrated that TAAR1 is upregulated in peripheral blood Cells Lines mononuclear cells (PBMC) and B cells following immune activation, and that subsequent • We performed Western blotting experiments to activation of TAAR1 by methamphetamine stimulates cAMP, similar to the function of measure levels of phospho-PKA and phospho- adenosine A2 receptors which are also present in immune cells and play a critical role in the PKC. immune response. Here, we are investigating the relationship between TAAR1 and the • We found that specific activators of PKA and adenosine A2 receptor at the level of cellular signaling and receptor dimerization. -

Adrenergic Agonist, Methoxamine, Reduces Exercise-Induced Asthma
Eur Respir J 1989, 2, 409-414 Pretreatment with an inhaled a 1-adrenergic agonist, methoxamine, reduces exercise-induced asthma A.T. Dinh Xuan, M. Chaussain, J. Regnard, A. Lockhart PretreatmenJ with an inhaled a,-adrenergic agonist. methoxamine, reduces Laboratoire de Physiologie, Faculte de Medecine eurcise-induced asthma. A.T. Dinh Xuan, M. Chaussain, J. Regnard, A. Cochin Port-Royal et Laboratoires d'Exploration Lockhart. Fonctionnelle Respiratoire, Hopitaux Cochin et ABSTRACT: In order to assess the role of the bronchial circulation in the Saint Vincent de Paul, 75014 Paris, France. pathogenes.ls of exercise-.induced asthma (EIA), we conducted a double Correspondence: Dr A.T. Dinh Xuan, Laboratoire blind, randomlzed study of the effects of pretreatment with an inhaled a - 1 d 'Explorations Fonctionnelles, Pavilion Potain, adrenergic agonist, methoxamine (Mx), l.n nine asthmatic teenagers with Hopital Cochin, 27 Rue du Faubourg Saint known r..:rA. Exercise consisted of S min cycle ergometry at a submaximal, Jacques, 75014 Paris, France. constant work-load, while the subjecfs breathed dry air at ambient tem ) Keywords: Airway oedema; a.,-adrenoceptors; perature. Forced expiratory volume in one second (FEV1 was measured at ba...eJine, JS min after pretreatment of either Mx or saline, and ser ially bronchial circulation; exercise-induced asthma; methoxamine. after exercise. Mx significantly reduced the exercise-induced fa ll of FEV 1 without modifying baseline FEV1 In five of the eight subJec t~ , had li ttle or no effect In three and caused an acute asthmatic attack in the remaining Received: June, 1988; accepted after revision December 29, 1988. subject. Mx has p<>tent constrictor effects on both bronchial and vascular smooth muscles through stimulation of postjunctlonal a 1-nd renoceptors. -

MDPV Bath Salts Test
One Step Methylenedioxypyrovalerone Drug of Abuse Test MATERIALS Analytical Sensitivity The cut-off concentration of the One Step Methylenedioxypyrovalerone Drug of Abuse Test is determined to be 1,000ng/mL. (Dip Card) Materials Provided: Test was run in 30 replicates with negative urine and standard control at ±25% cut-off and ±50% cut-off concentration levels. Test results ● Dip cards are summarized below. For Forensic Use Only ● Desiccants ● Package insert Test Result INTENDED USE Percent of Cut-off n Materials Required But Not Provided: Methylenedioxypyrovalerone Concentration in ng/mL The One Step Methylenedioxypyrovalerone Drug of Abuse Test is a lateral flow chromatographic immunoassay for the ● Specimen collection container Negative Positive qualitative detection of Methylenedioxypyrovalerone (MDPV) in human urine specimen at the cut-off level of 1,000ng/mL. This ● Disposable gloves 0% Cut-off assay is intended for forensic use only. ● Timer 30 30 0 This assay provides only a preliminary qualitative test result. A more specific confirmatory reference method, such as Liquid (No Drug Present) chromatogra -50% Cut-off phy tandem mass spectrometry (LC/MS/MS) or gas chromatography/mass spectrometry (GC/MS) must be use in INSTRUCTIONS FOR USE] 30 30 0 order to obtain a confirmed analytical result. (500ng/mL) 1) Remove the dip card from the foil pouch. -25% Cut-off 30 30 0 BACKGROUND 2) Remove the cap from the dip card. Label the device with patient or control identifications. (750ng/mL) 3) Immerse the absorbent tip into the urine sample for 5 seconds. Urine sample should not touch the plastic device. Cut-off ‘Bath salts’, a form of designer drugs, also promoted as ‘plant food’ or ‘research chemicals’, is sold mainly in head shops, on 4) Replace the cap over the absorbent tip and lay the dip card on a clean, flat, and non-absorptive surface. -

Certificate of Analysis
Print Date: Jan 3rd 2020 Certificate of Analysis www.tocris.com Product Name: EPPTB Catalog No.: 4518 Batch No.: 1 CAS Number: 1110781-88-8 IUPAC Name: N-(3-Ethoxyphenyl)-4-(1-pyrrolidinyl)-3-(trifluoromethyl)benzamide 1. PHYSICAL AND CHEMICAL PROPERTIES Batch Molecular Formula: C20H21F3N2O2 Batch Molecular Weight: 378.39 Physical Appearance: White solid Solubility: DMSO to 100 mM ethanol to 100 mM Storage: Store at +4°C Batch Molecular Structure: 2. ANALYTICAL DATA TLC: Rf = 0.2 (Ethyl acetate:Petroleum ether [4:1]) HPLC: Shows 99.9% purity 1H NMR: Consistent with structure Mass Spectrum: Consistent with structure Microanalysis: Carbon Hydrogen Nitrogen Theoretical 63.48 5.59 7.4 Found 63.37 5.53 7.44 Caution - Not Fully Tested • Research Use Only • Not For Human or Veterinary Use bio-techne.com North America China Europe Middle East Africa Rest of World [email protected] Tel: (800) 343 7475 [email protected] Tel: +44 (0)1235 529449 www.tocris.com/distributors [email protected] Tel: +86 (21) 52380373 Tel:+1 612 379 2956 Print Date: Jan 3rd 2020 Product Information www.tocris.com Product Name: EPPTB Catalog No.: 4518 Batch No.: 1 CAS Number: 1110781-88-8 IUPAC Name: N-(3-Ethoxyphenyl)-4-(1-pyrrolidinyl)-3-(trifluoromethyl)benzamide Description: Storage: Store at +4°C Trace amine 1 (TA ) receptor antagonist/inverse agonist; 1 Solubility & Usage Info: exhibits a higher potency at the mouse TA1 receptor than the rat DMSO to 100 mM and human TA150 receptors (IC values are 27.5, 4539 and 7487 ethanol to 100 mM nM, respectively).