Watada, M., Amd M. Itoh
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
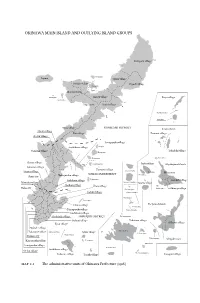
Okinawa Main Island and Outlying Island Groups
OKINAWA MAIN ISLAND AND OUTLYING ISLAND GROUPS Kunigami village Kourijima Iejima Ōgimi village Nakijin village Higashi village Yagajijima Ōjima Motobu town Minnajima Haneji village Iheya village Sesokojima Nago town Kushi village Gushikawajima Izenajima Onna village KUNIGAMI DISTRICT Kerama Islands Misato village Kin village Zamami village Goeku village Yonagusuku village Gushikawa village Ikeijima Yomitan village Miyagijima Tokashiki village Henzajima Ikemajima Chatan village Hamahigajima Irabu village Miyakojima Islands Ginowan village Katsuren village Kita Daitōjima Urasoe village Irabujima Hirara town NAKAGAMI DISTRICT Simojijima Shuri city Nakagusuku village Nishihara village Tsukenjima Gusukube village Mawashi village Minami Daitōjima Tarama village Haebaru village Ōzato village Kurimajima Naha city Oki Daitōjima Shimoji village Sashiki village Okinotorishima Uozurijima Kudakajima Chinen village Yaeyama Islands Kubajima Tamagusuku village Tono shirojima Gushikami village Kochinda village SHIMAJIRI DISTRICT Hatomamajima Mabuni village Taketomi village Kyan village Oōhama village Makabe village Iriomotejima Kumetorishima Takamine village Aguni village Kohamajima Kume Island Itoman city Taketomijima Ishigaki town Kanegusuku village Torishima Kuroshima Tomigusuku village Haterumajima Gushikawa village Oroku village Aragusukujima Nakazato village Tonaki village Yonaguni village Map 2.1 The administrative units of Okinawa Prefecture (1916) <UN> Chapter 2 The Okinawan War and the Comfort Stations: An Overview (1944–45) The sudden expansion -

Practical and Advanced Renewable Energy in Okinawa
The First International Workshop on Open Energy Systems (14-15 January 2014, OIST) Practical and Advanced Renewable Energy in Okinawa Dr. Jun-ichiro Giorgos TSUTSUMI Professor, Faculty of Engineering E-mail: [email protected] Contents of Presentation • Emission of Green House Gas from Energy • Ordinary Popular Natural Energy in Okinawa – Natural Energy (Photovoltaic System, Wind Turbine) – Recycle Energy (Waste Heat, Digested Gas, BDF) • Hawaii-Okinawa Clean Energy Partnership – Smart Grid System, OTEC, Energy Saving, People Exchange • Energy Research in University of the Ryukyus – Remote Control, Ocean Biomass, Solar Heater, Power Stabilizer • Smart Energy Projects by Okinawa Prefecture – Smart Energy Houses, Leveling System, Miyako Projects, etc. • Energy Projects in Miyako Island – Whole Island EMS, PV on Rented Roofs, Small EV • Remarks in Development of Renewable Energy Global Air Temperature and CO2 Concentration Industrial Revolu0on CO2 Emission Rate from Fossil Fuels Heating values and CO2 emission rates by combustion of various fossil fuels. CO2 emission: Coal > Oil > Gas. CO2 Emission Rate by Electric Power Companies before Fukushima “Adjusted rate” means the emission rates adjusted by the carbon credits of Kyoto mechanism. Mega Solar Fields in Okinawa (1) Fukuzato, Miyakojima (4,000kW) Okinawa Electric Power Co. (2) Abu, Nago (1,000kW) Okinawa Electric Co. (3) Ikehara, Okinawa (2,000kW) EcoLumiere LLC. Mega Solar Energy Field in Miyako Island Damages on wind turbines By typhoon 0314 (Maemi) New Type of Tiltable Wind -

United Nations Security Council
United Nations Security Council Chairs: Natalia Constantine Katie Hamlin Committee Topics: Topic 1: Nuclear Weapons in North Korea Topic 2: Syria Cessation of Hostilities Topic 3: Senkaku Islands Dispute 0 Chair Biographies Hello delegates! My name is Natalia Constantine and I’m in my senior year. I have been in MUN since 8th grade and this is my second time chairing, but my first time participating in Security Council. Besides MUN, I am on the tennis team, masque, and the school’s mathletics team. I am hoping for a fun and educational conference with resolutions passed, please do not hesitate in contacting me at [email protected] if you have any questions. See you all December 10th! Good day delegates! I am Katherine Hamlin, and I am a Junior this year at New Hartford High School. I have participated in MUN since I was in 8th grade and this will be my first time serving as a chair. Outside of MUN, I am also a distance swimmer for our school’s Varsity Swim team, a member of our French and Latin clubs, and a member of our Students for Justice and Equality club. I am greatly looking forward to serving as a chair on Security Council, and I hope that it will be an interesting and productive committee. If you have any questions, please contact me at [email protected] See you in December! Special Committee Notes Our simulation of the Security Council will be run Harvard Style. This means that there will be NO pre-written resolutions allowed in committee. -

KAKEHASHI Project Okinawa Program the 1 Slot Program Report
Japan’s Friendship Ties Program (USA) KAKEHASHI Project Okinawa Program the 1st slot Program Report 1. Program Overview Under the “KAKEHASHI Project” of Japan’s Friendship Ties Program, 42 high school students and 4 supervisors from the United States visited Japan from December 6th to December 13th, 2016 to participate in the program aimed at promoting their understanding of Japan with regard to Japanese politics, economy, society, culture, history, and foreign policy. Through lecture by ministry, observation of historical sites, school exchange, homestay, and other experiences, the participants enjoyed a wide range of opportunities to improve their understanding of Japan and shared their individual interests and experiences through SNS. Based on their findings and learning in Japan, each group of participants made a presentation in the final session and reported on the action plans to be taken after returning to their home country. 【Participating Countries and Number of Participants】 U.S.A. 46 Participants (A: Illinois University Laboratory High School, B: Fort Hayes Arts and Academic High School) 【Prefectures Visited】 Tokyo, Okinawa 2. Program Schedule December 6th (Tue) Arrival at Narita International Airport December 7th (Wed) [Orientation] [Lecture] North American Affairs Bureau, Ministry of Foreign Affairs “Japan’s Foreign Policy” [Historical Landmark] Imperial Palace Move to Okinawa December 8th (Thu) [Historical Facilities] Okinawa Prefectural Peace Memorial Museum, Peace Memorial Park [Historical Landmark] Shurijo Castle [Observation] Okinawa Prefectural Museum [Cultural Experience] Eisa dance 1 December 9th (Fri) [School Experience・Homestay] Okinawa Prefectural Naha Kokusai High School (Group A), Okinawa Prefectural Nago High School (Group B) December 10th (Sat) [Homestay] December 11th (Sun) [Homestay] Farewell Party [Workshop] December 12nd (Mon) Move to Tokyo [Reporting Session] December 13th (Tue) [Historical Landmark] Asakusa [Historical Landmark] Meiji Jingu Shrine Departure from Narita International Airport 3. -

Deconstructing Japan's Claim of Sovereignty Over the Diaoyu
Volume 10 | Issue 53 | Number 1 | Article ID 3877 | Dec 30, 2012 The Asia-Pacific Journal | Japan Focus Deconstructing Japan’s Claim of Sovereignty over the Diaoyu/Senkaku Islands 釣魚/尖閣に対する日本の統治権を脱構築 する Ivy Lee, Fang Ming “The near universal conviction in Japan with has shown effective control to be determinative which the islands today are declared an in a number of its rulings, a close scrutiny of ’integral part of Japan’s territory‘ is remarkable Japan’s effective possession/control reveals it to for its disingenuousness. These are islands have little resemblance to the effective unknown in Japan till the late 19th century possession/control in other adjudicated cases. (when they were identified from British naval As international law on territorial disputes, in references), not declared Japanese till 1895, theory and in practice, does not provide a not named till 1900, and that name not sound basis for its claim of sovereignty over the revealed publicly until 1950." GavanDiaoyu/Senkaku Islands, Japan will hopefully McCormack (2011)1 set aside its putative legal rights and, for the sake of peace and security in the region, start Abstract working with China toward a negotiated and mutually acceptable settlement. In this recent flare-up of the island dispute after Japan “purchased” three of theKeywords Diaoyu/Senkaku Islands, Japan reiterates its position that “the Senkaku Islands are an Diaoyu Dao, Diaoyutai, Senkakus, territorial inherent part of the territory of Japan, in light conflict, sovereignty, of historical facts and based upon international law.” This article evaluates Japan’s claims as I Introduction expressed in the “Basic View on the Sovereignty over the Senkaku Islands”A cluster of five uninhabited islets and three published on the website of the Ministry of rocky outcroppings lies on the edge of the East Foreign Affairs, Japan. -

Nansei Islands Biological Diversity Evaluation Project Report 1 Chapter 1
Introduction WWF Japan’s involvement with the Nansei Islands can be traced back to a request in 1982 by Prince Phillip, Duke of Edinburgh. The “World Conservation Strategy”, which was drafted at the time through a collaborative effort by the WWF’s network, the International Union for Conservation of Nature (IUCN), and the United Nations Environment Programme (UNEP), posed the notion that the problems affecting environments were problems that had global implications. Furthermore, the findings presented offered information on precious environments extant throughout the globe and where they were distributed, thereby providing an impetus for people to think about issues relevant to humankind’s harmonious existence with the rest of nature. One of the precious natural environments for Japan given in the “World Conservation Strategy” was the Nansei Islands. The Duke of Edinburgh, who was the President of the WWF at the time (now President Emeritus), naturally sought to promote acts of conservation by those who could see them through most effectively, i.e. pertinent conservation parties in the area, a mandate which naturally fell on the shoulders of WWF Japan with regard to nature conservation activities concerning the Nansei Islands. This marked the beginning of the Nansei Islands initiative of WWF Japan, and ever since, WWF Japan has not only consistently performed globally-relevant environmental studies of particular areas within the Nansei Islands during the 1980’s and 1990’s, but has put pressure on the national and local governments to use the findings of those studies in public policy. Unfortunately, like many other places throughout the world, the deterioration of the natural environments in the Nansei Islands has yet to stop. -

Higashi Village
We ask for your understanding Cape Hedo and cooperation for the environmental conservation funds. 58 Covered in spreading rich green subtropical forest, the northern part of 70 Okinawa's main island is called“Yanbaru.” Ferns and the broccoli-like 58 Itaji trees grow in abundance, and the moisture that wells up in between Kunigami Village Higashi Convenience Store (FamilyMart) Hentona Okinawa them forms clear streams that enrich the hilly land as they make their way Ie Island Ogimi Village towards the ocean. The rich forest is home to a number of animals that Kouri Island Prefecture cannot be found anywhere else on the planet, including natural monu- Okinawa Churaumi Aquarium Higashi Nakijin Village ments and endemic species such as the endangered Okinawa Rail, the (Ocean Expo Park) Genka Shioya Bay Village 9 Takae Okinawan Woodpecker and the Yanbaru Long-Armed Scarab Beetle, Minna Island Yagaji Island 331 Motobu Town 58 Taira making it a cradle of precious flora and fauna. 70 Miyagi Senaga Island Kawata Village With its endless and diverse vegetation, Yanbaru was selected as a 14 Arume Gesashi proposed world natural heritage site in December 2013. Nago City Living alongside this nature, the people of Yanbaru formed little settle- 58 331 ments hugging the coastline. It is said that in days gone by, lumber cut Kyoda I.C. 329 from the forest was passed from settlement to settlement, and carried to Shurijo Castle. Living together with the natural blessings from agriculture Futami Iriguchi Cape Manza and fishing, people's prayers are carried forward to the future even today Ginoza I.C. -

Militarization and Demilitarization of Okinawa As a Geostrategic “Keystone” Under the Japan-U.S
Militarization and Demilitarization of Okinawa As a Geostrategic “Keystone” under the Japan-U.S. Alliance August 10-12, 2013 International Geographical Union (IGU) 2013 Kyoto Regional Conference Commission on Political Geography Post-Conference Field Trip In Collaboration with Political Geography Research Group, Human Geographical Society of Japan and Okinawa Geographical Society Contents Organizers and Participants………………………………………………………………………….. p. 2 Co-organizers Assistants Supporting Organizations Informants Participants Time Schedule……………………………………………………………………………………….. p. 4 Route Maps……………………………………………………………………………………….…..p. 5 Naha Airport……………………………………………………………………………………….... p. 6 Domestic Flight Arrival Procedures Domestic Flight Departure Procedures Departing From Okinawa during a Typhoon Traveling to Okinawa during a Typhoon Accommodation………………………………………...…………………………………………..... p. 9 Deigo Hotel History of Deigo Hotel History of Okinawa (Ryukyu)………………………………………..………………………............. p. 11 From Ryukyu to Okinawa The Battle of Okinawa Postwar Occupation and Administration by the United States Post-Reversion U.S. Military Presence in Okinawa U.S. Military Bases in Okinawa…………………………………………………………………...… p. 14 Futenma Air Station Kadena Air Base Camp Schwab Camp Hansen Military Base Towns in Okinawa………………………………………………………...………….. p. 20 Political Economic Profile of Selected Base Towns Okinawa City (formerly Koza City) Chatan Town Yomitan Village Henoko, Nago City Kin Town What to do in Naha……………………………………………………………………………...… p. 31 1 Organizers -

Q&A on the Senkaku Islands
Q&A on the Senkaku Islands Q1: What is the basic view of the Government of Japan on the Senkaku Islands? A1: There is no doubt that the Senkaku Islands are clearly an inherent territory of Japan, in light of historical facts and based upon international law. Indeed, the Senkaku Islands are under the valid control of Japan. There exists no issue of territorial sovereignty to be resolved concerning the Senkaku Islands. Q2: What are the grounds for Japan's territorial sovereignty over the Senkaku Islands? A2: 1. From 1885 on, surveys of the Senkaku Islands had been thoroughly made by the Government of Japan by the agencies of Okinawa Prefecture and by way of other methods. Through these surveys, it was confirmed that the Senkaku Islands had been uninhabited and showed no trace of having been under the control of the Qing Dynasty of China. Based on this confirmation, the Government of Japan made a Cabinet Decision on 14 January 1895 to erect a marker on the Islands to formally incorporate the Senkaku Islands into the territory of Japan. These measures were carried out in accordance with the ways of duly acquiring territorial sovereignty under international law. 2. Since then, the Senkaku Islands have continuously remained as an integral part of the Nansei Shoto which are the territory of Japan. These islands were neither part of the island of Formosa nor part of the Pescadores Islands which were ceded to Japan from the Qing Dynasty of China in accordance with Article 2 of the Treaty of Shimonoseki which came into effect in May of 1895. -

Page 1 ACTA ARACHNOL., 27, Special Number), 1977. 337
ACTAARACHNOL.,27,Specialnumber),1977. 337 PreliminaryReportontheCaveSpider FaunaoftheRyukyuArchipelago By MatsueiSHIMOJANA FutenmaHighSchool,Iiutenma,GinowanCity,OkinawaPrefecture,Japan Synopsis SHIMOJANA,Matsuei(FutenmaHighSchool,Futenma,GinowanCity,OkinawaPre- fecture):PreliminaryreportonthecavespiderfaunaoftheRyukyuArchipelago.Actor arachnol.,27(Specialnumber):337-365(1977). AsurveyofthecavespiderfaunaintheRyukyuArchipelagowascarriedoutfrom1966 to1976.Fourtytwospeciesofthirtysixgenerabelongingtotwentythreefamilieswererecord- edfrommanylimestonecavesintheRyukyuIslands.Therepresentativecavespidersinthe RyukyuArchipelagoare〃 σ∫ゴ7σ〃σ10〃8ψ 姥 ρガ∫,ノ7αZ6ガ勿)!o〃6!σo々 勿 σ"σ6η ∫ガ3,の60667σ1σ 〃76σ♂σ andTetrablemmashimojanaietc.Amongofthem,SpeoceralaureatesandTetrablemma shimojanaiarewidelydistributedintheRyukyuChain.ThecavespiderfaunaoftheRyukyu ArchipelagoismuchdifferentfromtheJapaneseIslands. Introduction ThecavespiderfaunaoftheRyukyuIslandshasbeenreportedbyDr・T・ YAGINuMA(1962,1970),Dr.T.KoMATsu(1968,1972,1974)andthepresentauthor (1973),butthenumberofrecordedspeciesisfewandfragmentary・ BiospeleologicalsurveyoftheRyukyuArchipelagohavebeencarriedoutbythe author,andhehascollectedmanykindsofsubterraneananimalsfrommanylimestone caves. Thepresentpaperdealswiththespiderfaunadisclosedduringthesesurveys. Befbregoingfurtherintothesubjects,theauthorwishestoexpresshishearty thankstoDr.TakeoYAGINuMAofOhtemonGakuinUniversity,Osaka,Dr.Shun-ichi UENoofNationalScienceMuseum,Tokyo,Dr.SadaoIKEHARAofUniversityofthe Ryukyus,Okinawa,Dr.ToshihiroKoMATsuofMatsumotoDentalCollege,Nagano -

Onna Village Illustration Map
Onna Village Illustration Map Onna Sunset Coast Road This is the name given to the North-South 27 km (Ukaji to Nakama) Onna Village stretch of Prefectural Road 6 and National Route 58, which passes through Onna Village along the sea. Out of numerous recommen- Area:50.82k㎡ dations of a nickname from the public, this name was chosen to Population:11,089人 reflect the beautiful sunset view. The area with this designation was (As of January 2021) extended in the spring of 2014. The sea area within Onna Village is designated the Okinawa Seashore National Park and at dusk, the clear sky and waters are wrapped in a magical orange hue. → ❶ Manzamo (Prefectural Designated Scenic Beauty) ❶ Manzamo Tourist Facility ❷ Onna Village Seaside Park Nabee Beach The park features a flat meadow that extends to the edge This facility has souvenir shops, restaurants, an You can enjoy BBQ and marine sports throughout To Nago of a 20 m-high Ryukyu limestone cliff. The name is said exhibition space and an observatory. It is a the year at the beach. Protective jellyfish nets are Halekulani Okinawa [ Language] to originate from the words of the former king of Ryukyu, three-story building with a local specialty corner installed from April until the end of October. The UMITO PLAGE Shokei, who praised the place as being “a Mo (field) that on the first floor, restaurants and shops on the beach is located inside a bay so the sea is relatively The Atta Okinawa is big enough for 10,000 people (Man) to sit (za).” The second floor, and an observatory overlooking the calm, and you can enjoy swimming safely.The Hyatt Regency sunset is also beautiful and shouldn’t be missed. -

Corporate Guide.Indd
Corporate Guide Corporate Guide Published/March 2016 Business Planning Division, Planning & Research Department, The Okinawa Development Finance Corporation 1-2-26 Omoromachi, Naha-shi, Okinawa 900-8520 TEL.098-941-1740 FAX.098-941-1925 http://www.okinawakouko.go.jp/ ◎This pamphlet has been prepared using environmentally-friendly vegetable oil ink and recycled paper. CORPORATE GUIDE CONTENTS Overview of The Okinawa Development Finance Corporation Additional Materials Profile 02 History 20 Overview of Operations 03 Organization 21 Branches 22 Outline of Loans and Investment Systems Types of Funds 06 Industrial Development Loans 07 Small and Medium-sized Enterprise(SME) Loans 08 Micro Business Loans 09 Environmental Health Business Loans 10 Fuzhou Medical Service Loans 11 Taipei Primary Sector Loans 12 (Agriculture, Forestry, Fisheries) Housing Loans 13 Investments 14 Investments for the Creation of New Businesses 15 The Okinawa Development Finance Corporation Unique Funding Systems 16 Io-Torishima Island Okinawa Islands Iheya Island Izena Island Aguni Island Ie Island Okinawa Main Island Kume Island Senkaku Islands Taisho Island Kerama Islands Kuba Island Kitadaito Island Overview of The Okinawa Uotsuri Island Minamidaito Island Development Finance Corporation Daito Islands Irabu Island Miyako Island Sakishima Islands Profile ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 02 Yonaguni Island Tarama Island Miyako Islands Iriomote Island Ishigaki Island ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ Okidaito Island Overview of Operations