Genetic Analyses of Casuarinas Using ISSR and FISSR Markers
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Volume 5 Pt 3
Conservation Science W. Aust. 7 (3) : 583–592 (2010) Flora and vegetation of banded iron formations of the Yilgarn Craton: Mt Ida Greenstone Belt and Mt Hope RACHEL MEISSNER AND GAYNOR OWEN Science Division, Department of Environment and Conservation, PO Box 51, Wanneroo, Western Australia, 6946. Email: [email protected] ABSTRACT The Mount Ida greenstone belt is located within the Southern Cross Granite-Greenstone terrane (Wyche 2003) and is expressed as low ranges of banded ironstone formation. Recent increases in the value of iron ore have made the mining of banded iron formations more attractive. This paper describes the flora and vegetation on these ranges. A total eighty seven taxa, including one annual, were recorded from the ranges. One priority species (Atkins 2008), Calytrix erosipetala, and two range extensions were recorded. Hierarchical classification identified four plant communities on the ranges. The dominant floristic communities occurred on the slopes and crests (Community 2) and on the lower and colluvial slopes (Community 3). Upland communities were less fertile and there was a lower concentration of soil mobile elements than in the lowland communities. Currently, the Mt Ida Greenstone Belt is not within a conservation estate. Keywords: BIF, banded ironstone, floristic communities, ranges, Yilgarn INTRODUCTION The main range of the Mt Ida Greenstone Belt can be divided into two areas; the Mount Mason range in the Banded iron formations (BIF) are ancient sedimentary north extending 13km, and the Mt Ida Range, south of rocks formed in the Archaean period approximately 3.8 Mt Mason and extending c. 7km. The Mt Mason range is to 2.5 billion years ago. -

Clearing Permit Decision Report
Clearing Permit Decision Report 1. Application details 1.1. Permit application details Permit application No.: 8786/1 Permit type: Purpose Permit 1.2. Proponent details Proponent’s name: Mid-West Tungsten Pty Ltd 1.3. Property details Property: Mining Lease 59/386 Mining Lease 59/387 Mining Lease 59/425 Miscellaneous Licence 59/161 Miscellaneous Licence 59/162 Local Government Area: Shire of Perenjori Colloquial name: Mt Mulgine Project 1.4. Application Clearing Area (ha) No. Trees Method of Clearing For the purpose of: 201.8 Mechanical Removal Mineral Production and Associated Activities 1.5. Decision on application Decision on Permit Application: Refuse Decision Date: 24 September 2020 Reasons for Decision The clearing permit application was received on 08 January 2020 and has been assessed against the clearing principles, planning instruments, and other matters in accordance with section 51O of the Environmental Protection Act 1986 (EP Act). The initial assessment of the application determined that the proposed clearing would likely result in significant impacts to conservation significant flora and fauna, and was therefore unlikely to be considered acceptable. The applicant was given the opportunity to provide additional information and/or modify their clearing proposal in order to reduce the environmental impacts. In response, Mid-West Tungsten Pty Ltd provided additional information and biological survey reports. The additional information and survey reports were taken into consideration when making the decision on this application. -

Charles Darwin, Kadji Kadji, Karara, Lochada Reserves WA
BUSH BLITZ SPECIES DISCOVERY PROGRAM Charles Darwin Reserve WA 3–9 May · 14–25 September · 7–18 December 2009 Kadji Kadji, Karara, Lochada Reserves WA 14–25 September · 7–18 December 2009 What is Contents Bush Blitz? Bush Blitz is a four-year, What is Bush Blitz 2 multi-million dollar Summary 3 partnership between the Abbreviations 3 Australian Government, Introduction 4 BHP Billiton, and Earthwatch Reserves Overview 5 Australia to document plants Methods 8 and animals in selected properties across Australia’s Results 10 National Reserve System. Discussion 12 Appendix A: Species Lists 15 Fauna 16 This innovative partnership Vertebrates 16 harnesses the expertise of many Invertebrates 25 of Australia’s top scientists from Flora 48 museums, herbaria, universities, Appendix B: Rare and Threatened Species 79 and other institutions and Fauna 80 organisations across the country. Flora 81 Appendix C: Exotic and Pest Species 83 Fauna 84 Flora 85 2 Bush Blitz survey report Summary Bush Blitz fieldwork was conducted at four National Reserve System properties in the Western Australian Avon Wheatbelt and Yalgoo Bioregions during 2009. This included a pilot study Abbreviations at Charles Darwin Reserve and a longer study of Charles Darwin, Kadji Kadji, Lochada and Karara reserves. Results include 651 species added to those known across the reserves and the discovery of 35 putative species new to science. The majority of ANHAT these new species occur within the heteroptera (plant bugs) and Australian Natural Heritage Assessment lepidoptera (butterflies and moths) taxonomic groups. Tool Malleefowl (Leipoa ocellata), listed as vulnerable under the EPBC Act federal Environmental Protection and Biodiversity Conservation Environment Protection and Biodiversity Act 1999 (EPBC Act), were observed on Charles Darwin Reserve. -
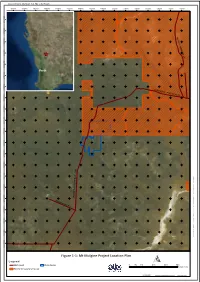
Tungsten Mining NL Mt Mulgine Project EPA REFERRAL
Document Name: 20170125_TUN_Fig1-1_Biol Report 482000 484000 486000 488000 490000 492000 494000 496000 498000 500000 502000 504000 506000 508000 510000 512000 0 0 0 2 9 7 6 0 0 0 0 9 7 6 Y A 0 L 0 G 0 O 8 8 O 7 6 - N I N G 0 0 H 0 A 6 N 8 7 R 6 ^_ D 0 0 0 4 8 7 6 Perth 0 0 0 2 8 7 6 0 0 0 0 8 7 6 W ARRIE DAR - PERE 0 NJO 0 RI RD 0 8 7 7 6 0 0 0 6 7 7 6 0 0 0 4 7 7 6 0 0 0 2 7 7 6 0 0 0 0 7 7 6 0 0 0 8 6 7 6 0 0 0 6 6 7 6 D R E 0 0 N I \ 0 \ M 4 M A 6 I N 7 R S 6 E E R V P E R P - P C O \ 0 s e C 0 r v 0 e R r 2 s t 6 A o r 7 a D g 6 e E \ I A P R M R G I A S 0 a 0 n W d 0 M 0 a 6 p 7 p i n 6 B O g O \ N 0 E 3 R _ O C N l i G e n 0 R D t \ 0 T U 0 N 8 \ 0 5 2 7 _ G 6 I S M a p s \ 2 0 0 1 0 7 0 0 1 6 2 5 5 _ T 7 U 6 N _ F i g 1 - 1 _ B 0 i o 0 l R 0 e 4 p o 5 r t 7 . -

Rangelands, Western Australia
Biodiversity Summary for NRM Regions Species List What is the summary for and where does it come from? This list has been produced by the Department of Sustainability, Environment, Water, Population and Communities (SEWPC) for the Natural Resource Management Spatial Information System. The list was produced using the AustralianAustralian Natural Natural Heritage Heritage Assessment Assessment Tool Tool (ANHAT), which analyses data from a range of plant and animal surveys and collections from across Australia to automatically generate a report for each NRM region. Data sources (Appendix 2) include national and state herbaria, museums, state governments, CSIRO, Birds Australia and a range of surveys conducted by or for DEWHA. For each family of plant and animal covered by ANHAT (Appendix 1), this document gives the number of species in the country and how many of them are found in the region. It also identifies species listed as Vulnerable, Critically Endangered, Endangered or Conservation Dependent under the EPBC Act. A biodiversity summary for this region is also available. For more information please see: www.environment.gov.au/heritage/anhat/index.html Limitations • ANHAT currently contains information on the distribution of over 30,000 Australian taxa. This includes all mammals, birds, reptiles, frogs and fish, 137 families of vascular plants (over 15,000 species) and a range of invertebrate groups. Groups notnot yet yet covered covered in inANHAT ANHAT are notnot included included in in the the list. list. • The data used come from authoritative sources, but they are not perfect. All species names have been confirmed as valid species names, but it is not possible to confirm all species locations. -

Declared Rare and Poorly Known Flora in the Geraldton District
WESTERN AUSTRALIAN WILDLIFE MANAGEMENT PROGRAM NO. 26 Declared Rare and Poorly Known Flora in the Geraldton District by Susan J. Patrick 2001 Department of Conservation and Land Management Locked Bag 104, Bentley Delivery Centre WA 6983 1 Department of Conservation and Land Management Locked Bag 104, Bentley Delivery Centre WA 6983 Department of Conservation and Land Management, Western Australia 2001 ISSN 0816-9713 Cover illustration: Verticordia spicata subsp. squamosa by Margaret Pieroni Editors ..........................................................................................................Angie Walker and Jill Pryde Page preparation ..................................................................................................................Angie Walker Maps ..................................................................................................... CALM Land Information Branch 2 FOREWORD Western Australian Wildlife Management Programs are a series of publications produced by the Department of Conservation and Land Management (CALM). The programs are prepared in addition to Regional Management Plans to provide detailed information and guidance for the management and protection of certain exploited or threatened species (e.g. Kangaroos, Noisy Scrub-bird and the Rose Mallee). This program provides a brief description of the appearance, distribution, habitat and conservation status of flora declared as rare under the Western Australian Wildlife Conservation Act (Threatened Flora) and poorly known flora (Priority -

Species List
Biodiversity Summary for NRM Regions Species List What is the summary for and where does it come from? This list has been produced by the Department of Sustainability, Environment, Water, Population and Communities (SEWPC) for the Natural Resource Management Spatial Information System. The list was produced using the AustralianAustralian Natural Natural Heritage Heritage Assessment Assessment Tool Tool (ANHAT), which analyses data from a range of plant and animal surveys and collections from across Australia to automatically generate a report for each NRM region. Data sources (Appendix 2) include national and state herbaria, museums, state governments, CSIRO, Birds Australia and a range of surveys conducted by or for DEWHA. For each family of plant and animal covered by ANHAT (Appendix 1), this document gives the number of species in the country and how many of them are found in the region. It also identifies species listed as Vulnerable, Critically Endangered, Endangered or Conservation Dependent under the EPBC Act. A biodiversity summary for this region is also available. For more information please see: www.environment.gov.au/heritage/anhat/index.html Limitations • ANHAT currently contains information on the distribution of over 30,000 Australian taxa. This includes all mammals, birds, reptiles, frogs and fish, 137 families of vascular plants (over 15,000 species) and a range of invertebrate groups. Groups notnot yet yet covered covered in inANHAT ANHAT are notnot included included in in the the list. list. • The data used come from authoritative sources, but they are not perfect. All species names have been confirmed as valid species names, but it is not possible to confirm all species locations. -

Flora and Vegetation of the Eastern Goldfields Ranges: Part 7
Journal of the Royal Society of Western Australia, 87:49–62, 2004 Flora and vegetation of the Eastern Goldfields Ranges: Part 7. Middle and South Ironcap, Digger Rock and Hatter Hill N Gibson Science Division, Department of Conservation and Land Management, Wildlife Research Centre, PO Box 51 Wanneroo WA 6065 [email protected] (Manuscript received July 2003; accepted June 2004) Abstract A study of the flora and plant communities of part of Forrestania greenstone belt between Middle Ironcap and Hatter Hill (some 80 km ESE of Hyden), recorded a total flora of 345 taxa of which 342 were native and three were introduced. Three species of threatened flora and 29 taxa being considered for listing were found. Ten species are considered to be endemic to the range and a further eight species are restricted to similar landforms within 100 km of the range. A new species of Stenanthemum is only known from two populations. Despite considerable mining and exploration activity in the area, the flora and vegetation remain poorly known. Thirty-eight quadrats were established along the range system and data from these quadrats were used to define four community types. Differences in these community types were strongly related to edaphic gradients. Very little of the Forrestania vegetation system is reserved and the results of this survey support recommendations for the establishment of nature reserves to conserve this vegetation system. Keywords: flora, vegetation, Goldfields, Ironcap, Hatter Hill, Digger Rock, Western Australia, greenstone Introduction wildfire in summer of 1993 that consumed almost all vegetation between Mt Holland and Middle Ironcap. -

Charles Darwin Reserve Project ABRS Final Report Plants
Biodiversity survey pilot project at Charles Darwin Reserve, Western Australia Vascular Plants Report to Australian Biological Resources Study, Department of the Environment, Water, Heritage and the Arts, Canberra Terry D. Macfarlane Western Australian Herbarium, Science Division, Department of Environment and Conservation, Western Australia. 31 August 2009 Cover picture: View of York gum ( Eucalyptus loxophleba ) woodland below a greenstone ridge, with extensive Acacia shrubland in the distance, northern Charles Darwin Reserve, May 2009. (Photo T.D. Macfarlane). 2 Biodiversity survey pilot project at Charles Darwin Reserve, Western Australia Terry D. Macfarlane Western Australian Herbarium, Science Division, Department of Environment and Conservation, Western Australia. Address: DEC, Locked Bag 2, Manjimup WA 6258 Project aim This project was conceived to carry out a biological survey of a reserve forming part of the National Reserve System, to provide biodiversity information for reserve management and to contribute to taxonomic knowledge including description of new species as appropriate. Survey structure A team of scientists specialising in different organism groups each with a team of Earthwatch volunteer assistants, driver and vehicle, carried out the survey following individually devised field programs, over the 1 week period 5-9 May 2009. Science teams Five science teams, each led by one of the scientists, specialised on the following organism groups: Plants, Insects (Lepidoptera), Insects (Heteroptera), Vertebrates , Arachnids. Plants were collected by the Plants team and also by some of the animal scientists, particularly Celia Symonds, in order to accurately document the insect host plants. These additional collections were identified along with the other flora vouchers, and contribute to the flora results reported here. -

Pages 583–592
Conservation Science W. Aust. 7 (3) : 583–592 (2010) Flora and vegetation of banded iron formations of the Yilgarn Craton: Mt Ida Greenstone Belt and Mt Hope RACHEL MEISSNER AND GAYNOR OWEN Science Division, Department of Environment and Conservation, PO Box 51, Wanneroo, Western Australia, 6946. Email: [email protected] ABSTRACT The Mount Ida greenstone belt is located within the Southern Cross Granite-Greenstone terrane (Wyche 2003) and is expressed as low ranges of banded ironstone formation. Recent increases in the value of iron ore have made the mining of banded iron formations more attractive. This paper describes the flora and vegetation on these ranges. A total eighty seven taxa, including one annual, were recorded from the ranges. One priority species, Calytrix erosipetala, and two range extensions were recorded. Hierarchical classification identified four plant communities on the ranges. The dominant floristic communities occurred on the slopes and crests (Community 2) and on the lower and colluvial slopes (Community 3). Upland communities were less fertile and there was a lower concentration of soil mobile elements than in the lowland communities. Currently, the Mt Ida Greenstone Belt is not within a conservation estate. Keywords: BIF, banded ironstone, floristic communities, ranges, Yilgarn INTRODUCTION The main range of the Mt Ida Greenstone Belt can be divided into two areas; the Mount Mason range in the Banded iron formations (BIF) are ancient sedimentary north extending 13km, and the Mt Ida Range, south of rocks formed in the Archaean period approximately 3.8 Mt Mason and extending c. 7km. The Mt Mason range is to 2.5 billion years ago. -

A Summary of the Published Data on Host Plants and Morphology of Immature Stages of Australian Jewel Beetles (Coleoptera: Buprestidae), with Additional New Records
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Center for Systematic Entomology, Gainesville, Insecta Mundi Florida 3-22-2013 A summary of the published data on host plants and morphology of immature stages of Australian jewel beetles (Coleoptera: Buprestidae), with additional new records C. L. Bellamy California Department of Food and Agriculture, [email protected] G. A. Williams Australian Museum, [email protected] J. Hasenpusch Australian Insect Farm, [email protected] A. Sundholm Sydney, Australia, [email protected] Follow this and additional works at: https://digitalcommons.unl.edu/insectamundi Bellamy, C. L.; Williams, G. A.; Hasenpusch, J.; and Sundholm, A., "A summary of the published data on host plants and morphology of immature stages of Australian jewel beetles (Coleoptera: Buprestidae), with additional new records" (2013). Insecta Mundi. 798. https://digitalcommons.unl.edu/insectamundi/798 This Article is brought to you for free and open access by the Center for Systematic Entomology, Gainesville, Florida at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Insecta Mundi by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. INSECTA MUNDI A Journal of World Insect Systematics 0293 A summary of the published data on host plants and morphology of immature stages of Australian jewel beetles (Coleoptera: Buprestidae), with additional new records C. L. Bellamy G. A. Williams J. Hasenpusch A. Sundholm CENTER FOR SYSTEMATIC ENTOMOLOGY, INC., Gainesville, FL Cover Photo. Calodema plebeia Jordan and several Metaxymorpha gloriosa Blackburn on the flowers of the proteaceous Buckinghamia celcissima F. Muell. in the lowland mesophyll vine forest at Polly Creek, Garradunga near Innisfail in northeastern Queensland. -

Taimeselts Fagales Süstemaatika Ja Levik Maailmas
Tartu Ülikool Loodus- ja tehnoloogiateaduskond Ökoloogia ja Maateaduste Instituut Botaanika osakond Hanna Hirve TAIMESELTS FAGALES SÜSTEMAATIKA JA LEVIK MAAILMAS Bakalaureusetöö Juhendaja: professor Urmas Kõljalg Tartu 2014 Sisukord Sisukord ............................................................................................................................ 2 Sissejuhatus ...................................................................................................................... 4 1. Taimeseltsist Fagales üldiselt ................................................................................... 5 2. Takson Betulaceae ................................................................................................... 7 2.1 Iseloomustus ja levik ......................................................................................... 7 2.2 Morfoloogilised tunnused .................................................................................. 8 2.3 Fülogenees ......................................................................................................... 9 2.4 Tähtsus ............................................................................................................... 9 3. Takson Casuarinaceae ............................................................................................ 10 3.1 Iseloomustus ja levik ....................................................................................... 10 3.2 Morfoloogilised tunnused ...............................................................................