A New Genus and Species of Buteonine Hawk from Quaternary Deposits in Bermuda (Aves: Accipitridae)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Disaggregation of Bird Families Listed on Cms Appendix Ii
Convention on the Conservation of Migratory Species of Wild Animals 2nd Meeting of the Sessional Committee of the CMS Scientific Council (ScC-SC2) Bonn, Germany, 10 – 14 July 2017 UNEP/CMS/ScC-SC2/Inf.3 DISAGGREGATION OF BIRD FAMILIES LISTED ON CMS APPENDIX II (Prepared by the Appointed Councillors for Birds) Summary: The first meeting of the Sessional Committee of the Scientific Council identified the adoption of a new standard reference for avian taxonomy as an opportunity to disaggregate the higher-level taxa listed on Appendix II and to identify those that are considered to be migratory species and that have an unfavourable conservation status. The current paper presents an initial analysis of the higher-level disaggregation using the Handbook of the Birds of the World/BirdLife International Illustrated Checklist of the Birds of the World Volumes 1 and 2 taxonomy, and identifies the challenges in completing the analysis to identify all of the migratory species and the corresponding Range States. The document has been prepared by the COP Appointed Scientific Councilors for Birds. This is a supplementary paper to COP document UNEP/CMS/COP12/Doc.25.3 on Taxonomy and Nomenclature UNEP/CMS/ScC-Sc2/Inf.3 DISAGGREGATION OF BIRD FAMILIES LISTED ON CMS APPENDIX II 1. Through Resolution 11.19, the Conference of Parties adopted as the standard reference for bird taxonomy and nomenclature for Non-Passerine species the Handbook of the Birds of the World/BirdLife International Illustrated Checklist of the Birds of the World, Volume 1: Non-Passerines, by Josep del Hoyo and Nigel J. Collar (2014); 2. -

Chromosome Painting in Three Species of Buteoninae: a Cytogenetic Signature Reinforces the Monophyly of South American Species
Chromosome Painting in Three Species of Buteoninae: A Cytogenetic Signature Reinforces the Monophyly of South American Species Edivaldo Herculano C. de Oliveira1,2,3*, Marcella Mergulha˜o Tagliarini4, Michelly S. dos Santos5, Patricia C. M. O’Brien3, Malcolm A. Ferguson-Smith3 1 Laborato´rio de Cultura de Tecidos e Citogene´tica, SAMAM, Instituto Evandro Chagas, Ananindeua, PA, Brazil, 2 Faculdade de Cieˆncias Exatas e Naturais, ICEN, Universidade Federal do Para´, Bele´m, PA, Brazil, 3 Cambridge Resource Centre for Comparative Genomics, Cambridge, United Kingdom, 4 Programa de Po´s Graduac¸a˜oem Neurocieˆncias e Biologia Celular, ICB, Universidade Federal do Para´, Bele´m, PA, Brazil, 5 PIBIC – Universidade Federal do Para´, Bele´m, PA, Brazil Abstract Buteoninae (Falconiformes, Accipitridae) consist of the widely distributed genus Buteo, and several closely related species in a group called ‘‘sub-buteonine hawks’’, such as Buteogallus, Parabuteo, Asturina, Leucopternis and Busarellus, with unsolved phylogenetic relationships. Diploid number ranges between 2n = 66 and 2n = 68. Only one species, L. albicollis had its karyotype analyzed by molecular cytogenetics. The aim of this study was to present chromosomal analysis of three species of Buteoninae: Rupornis magnirostris, Asturina nitida and Buteogallus meridionallis using fluorescence in situ hybridization (FISH) experiments with telomeric and rDNA probes, as well as whole chromosome probes derived from Gallus gallus and Leucopternis albicollis. The three species analyzed herein showed similar karyotypes, with 2n = 68. Telomeric probes showed some interstitial telomeric sequences, which could be resulted by fusion processes occurred in the chromosomal evolution of the group, including the one found in the tassociation GGA1p/GGA6. -

Tinamiformes – Falconiformes
LIST OF THE 2,008 BIRD SPECIES (WITH SCIENTIFIC AND ENGLISH NAMES) KNOWN FROM THE A.O.U. CHECK-LIST AREA. Notes: "(A)" = accidental/casualin A.O.U. area; "(H)" -- recordedin A.O.U. area only from Hawaii; "(I)" = introducedinto A.O.U. area; "(N)" = has not bred in A.O.U. area but occursregularly as nonbreedingvisitor; "?" precedingname = extinct. TINAMIFORMES TINAMIDAE Tinamus major Great Tinamou. Nothocercusbonapartei Highland Tinamou. Crypturellus soui Little Tinamou. Crypturelluscinnamomeus Thicket Tinamou. Crypturellusboucardi Slaty-breastedTinamou. Crypturellus kerriae Choco Tinamou. GAVIIFORMES GAVIIDAE Gavia stellata Red-throated Loon. Gavia arctica Arctic Loon. Gavia pacifica Pacific Loon. Gavia immer Common Loon. Gavia adamsii Yellow-billed Loon. PODICIPEDIFORMES PODICIPEDIDAE Tachybaptusdominicus Least Grebe. Podilymbuspodiceps Pied-billed Grebe. ?Podilymbusgigas Atitlan Grebe. Podicepsauritus Horned Grebe. Podicepsgrisegena Red-neckedGrebe. Podicepsnigricollis Eared Grebe. Aechmophorusoccidentalis Western Grebe. Aechmophorusclarkii Clark's Grebe. PROCELLARIIFORMES DIOMEDEIDAE Thalassarchechlororhynchos Yellow-nosed Albatross. (A) Thalassarchecauta Shy Albatross.(A) Thalassarchemelanophris Black-browed Albatross. (A) Phoebetriapalpebrata Light-mantled Albatross. (A) Diomedea exulans WanderingAlbatross. (A) Phoebastriaimmutabilis Laysan Albatross. Phoebastrianigripes Black-lootedAlbatross. Phoebastriaalbatrus Short-tailedAlbatross. (N) PROCELLARIIDAE Fulmarus glacialis Northern Fulmar. Pterodroma neglecta KermadecPetrel. (A) Pterodroma -
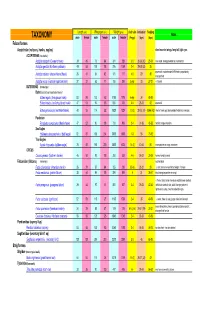
Avian Taxonomy
Length (cm) Wing span (cm) Weight (gms) cluch size incubation fledging Notes TAXONOMY male female male female male female (# eggs) (days) (days) Falconiformes Accipitridae (vultures, hawks, eagles) short rounded wings; long tail; light eyes ACCIPITRINAE (true hawks) Accipiter cooperii (Cooper's hawk) 39 45 73 84 341 528 3-5 30-36 (30) 25-34 crow sized; strongly banded tail; rounded tail Accipiter gentilis (Northern goshawk) 49 58 101 108 816 1059 2-4 28-38 (33) 35 square tail; most abundant NAM hawk; proportionaly 26 31 54 62 101 177 4-5 29 30 Accipiter striatus (sharp-shinned hawk) strongest foot Accipiter nisus (Eurasian sparrowhawk ) 37 37 62 77 150 290 5-Apr 33 27-31 Musket BUTEONINAE (broadwings) Buteo (buzzards or broad tailed hawks) Buteo regalis (ferruginous hawk) 53 59 132 143 1180 1578 6-Apr 34 45-50 Buteo lineatus (red-shouldered hawk) 47 53 96 105 550 700 3-4 28-33 42 square tail Buteo jamaicensis (red-tailed hawk) 48 55 114 122 1028 1224 1-3 (3) 28-35 (34) 42-46 (42) (Harlan' hawk spp); dark patagial featehres: immature; Parabuteo Parabuteo cuncincutus (Harris hawk) 47 52 90 108 710 890 2-4 31-36 45-50 reddish orange shoulders Sea Eagles Haliaeetus leucocephalus (bald eagle) 82 87 185 244 3000 6300 1-3 35 70-92 True Eagles Aquila chrysaetos (golden eagle) 78 82 185 220 3000 6125 1-4 (2) 40-45 50 white patches on wings: immature; CIRCUS Circus cyaneus (Northern harrier) 46 50 93 108 350 530 4-6 26-32 30-35 hovers; hunts by sound Falconidae (falcons) (longwings) notched beak Falco columbarius (American merlin) 26 29 57 64 -

Estimations Relative to Birds of Prey in Captivity in the United States of America
ESTIMATIONS RELATIVE TO BIRDS OF PREY IN CAPTIVITY IN THE UNITED STATES OF AMERICA by Roger Thacker Department of Animal Laboratories The Ohio State University Columbus, Ohio 43210 Introduction. Counts relating to birds of prey in captivity have been accomplished in some European countries; how- ever, to the knowledge of this author no such information is available in the United States of America. The following paper consistsof data related to this subject collected during 1969-1970 from surveys carried out in many different direc- tions within this country. Methods. In an attempt to obtain as clear a picture as pos- sible, counts were divided into specific areas: Research, Zoo- logical, Falconry, and Pet Holders. It became obvious as the project advanced that in some casesthere was overlap from one area to another; an example of this being a falconer working with a bird both for falconry and research purposes. In some instances such as this, the author has used his own judgment in placing birds in specific categories; in other in- stances received information has been used for this purpose. It has also become clear during this project that a count of "pets" is very difficult to obtain. Lack of interest, non-coop- eration, or no available information from animal sales firms makes the task very difficult, as unfortunately, to obtain a clear dispersal picture it is from such sourcesthat informa- tion must be gleaned. However, data related to the importa- tion of birds' of prey as recorded by the Bureau of Sport Fisheries and Wildlife is included, and it is felt some observa- tions can be made from these figures. -

SMCSP & SMCSN Wildlife List.Xlsx
Appendix C: Wildlife list for Six Mile Cypress Slough and Six Mile Cypress North Preserves Designated Status Scientific Name Common Name FWC FWS FNAI MAMMALS Family: Didelphidae (opossums) Didelphis virginiana Virginia opossum Family: Dasypodidae (armadillos) Dasypus novemcinctus nine-banded armadillo * Family: Sciuridae (squirrels and their allies) Sciurus carolinensis eastern gray squirrel Sciurus niger avicennia Big Cypress fox squirrel T G5T2/S2 Family: Muridae (mice and rats) Peromyscus gossypinus cotton mouse Oryzomys palustris marsh rice rat Sigmodon hispidus hispid cotton rat Family: Leporidae (rabbits and hares) Sylvilagus palustris marsh rabbit Sylvilagus floridanus eastern cottontail Family: Talpidae (moles) Scalopus aquaticus eastern mole Family: Felidae (cats) Puma concolor coryi Florida panther E E G5T1/S1 Lynx rufus bobcat Felis silvestris domestic cat * Family: Canidae (wolves and foxes) Urocyon cinereoargenteus common gray fox Family: Ursidae (bears) Ursus americanus floridanus Florida black bear T G5T2/S2 Family: Procyonidae (raccoons) Procyon lotor raccoon Family: Mephitidae (skunks) Spilogale putorius eastern spotted skunk Mephitis mephitis striped skunk Family: Mustelidae (weasels, otters and relatives) Lutra canadensis northern river otter Family: Suidae (old world swine) Sus scrofa feral hog * Family: Cervidae (deer) Odocoileus virginianus white-tailed deer BIRDS Family: Anatidae (swans, geese and ducks) Subfamily: Anatinae (dabbling ducks) Dendrocygna autumnalis black-bellied whistling duck Cairina moschata muscovy -

Journal of Avian Biology JAV-00869 Wang, N
Journal of Avian Biology JAV-00869 Wang, N. and Kimball, R. T. 2016. Re-evaluating the distribution of cooperative breeding in birds: is it tightly linked with altriciality? – J. Avian Biol. doi: 10.1111/jav.00869 Supplementary material Appendix 1. Table A1. The characteristics of the 9993 species based on Jetz et al. (2012) Order Species Criteria1 Developmental K K+S K+S+I LB Mode ACCIPITRIFORMES Accipiter albogularis 0 0 0 0 1 ACCIPITRIFORMES Accipiter badius 0 0 0 0 1 ACCIPITRIFORMES Accipiter bicolor 0 0 0 0 1 ACCIPITRIFORMES Accipiter brachyurus 0 0 0 0 1 ACCIPITRIFORMES Accipiter brevipes 0 0 0 0 1 ACCIPITRIFORMES Accipiter butleri 0 0 0 0 1 ACCIPITRIFORMES Accipiter castanilius 0 0 0 0 1 ACCIPITRIFORMES Accipiter chilensis 0 0 0 0 1 ACCIPITRIFORMES Accipiter chionogaster 0 0 0 0 1 ACCIPITRIFORMES Accipiter cirrocephalus 0 0 0 0 1 ACCIPITRIFORMES Accipiter collaris 0 0 0 0 1 ACCIPITRIFORMES Accipiter cooperii 0 0 0 0 1 ACCIPITRIFORMES Accipiter erythrauchen 0 0 0 0 1 ACCIPITRIFORMES Accipiter erythronemius 0 0 0 0 1 ACCIPITRIFORMES Accipiter erythropus 0 0 0 0 1 ACCIPITRIFORMES Accipiter fasciatus 0 0 0 0 1 ACCIPITRIFORMES Accipiter francesiae 0 0 0 0 1 ACCIPITRIFORMES Accipiter gentilis 0 0 0 0 1 ACCIPITRIFORMES Accipiter griseiceps 0 0 0 0 1 ACCIPITRIFORMES Accipiter gularis 0 0 0 0 1 ACCIPITRIFORMES Accipiter gundlachi 0 0 0 0 1 ACCIPITRIFORMES Accipiter haplochrous 0 0 0 0 1 ACCIPITRIFORMES Accipiter henicogrammus 0 0 0 0 1 ACCIPITRIFORMES Accipiter henstii 0 0 0 0 1 ACCIPITRIFORMES Accipiter imitator 0 0 0 0 1 ACCIPITRIFORMES -

Breeding Biology and Nestling Diet of the Great Black&Hyphen;Hawk
SHORT COMMUNICATIONS j. RaptorRes. 32(2):175-177 ¸ 1998 The Raptor Research Foundation, Inc. BREEDING BIOLOGY AND NESTLING DIET OF THE GREAT BLACK-HAWK NATHANIEL E. SEAVY 17142 LemoloShr. Dr. N.E., Poulsbo, WA 98370 U.S.A. RtCH?d•D P. GERHARDT 341 N.E. Chestnut St., Madras, OR 97741 U.S.A. KEY WORDS: GreatBlack-Hawk; Buteogallus urubitinga; Observationsof courtship behavior or of hawkscarry- breedingbiology; diet;, Petgn; Guatemala. ing nest material or prey led to the eventual location of nests.After they were found, nests were checked every 2-3 d to record nesting phenology.During all yearswe The Great Black-Hawk (Buteogallusurubitinga) rang- recorded nest size (diameter and depth) and situation, es from Mexico south to eastern Bolivia, Paraguayand and described nest trees. Observationsof prey deliveries to nests were made with binoculars from observation northern Argentina, inhabiting coastal lowlands and foothills (Brown and Amadon 1968). The few accounts platforms constructed in trees about 35 m from nests. We climbed to nestsweekly to weigh and measure nest- describing its breeding biology have been brief and at lings in 1991; in 1993 and 1994 we avoided climbing to times contradictory(Grossman and Hamlet 1964, Smithe nests,except to verify someclutch sizes,until after fledg- 1966, Brown and Amadon 1968, ffrench 1976, Mader ing. Additional information on clutch size, nesting phe- 1981). Based primarily on isolated observationsof hunt- nology and nestswas obtained from egg-setdata records ing and prey remains collected beneath roosts, a wide from published accounts,the Western Foundation of Ver- variety of prey items has been recorded, including inver- tebrate Zoology (WFVZ), and the Delaware Museum of tebrates,fish, frogs, reptiles,birds, mammalsand carrion Natural History (DMNH). -

Breeding Ecology and Behavior of the Hawaiian Hawk ’
The Condor 100:654-662 01 he‘ Cooper Omitholog~cal Society 1998 BREEDING ECOLOGY AND BEHAVIOR OF THE HAWAIIAN HAWK ’ CURTICE R. GRIFFIN*, PETER W. C. PATON~ AND THOMAS S. BASKEM Schoolof Forestry, Fisheries, and Wildlife, Universityof Missouri-Columbia,Columbia, MO 65211, e-mail: [email protected] Abstract. We studied the ecology of the endangeredHawaiian Hawk (Buteo solitarius) Downloaded from https://academic.oup.com/condor/article/100/4/654/5126040 by guest on 30 September 2021 on the island of Hawaii for three breeding seasons.Their breeding strategy is a prime example of a K-selected speciescharacteristic of many birds in tropical environments:clutch size was one and brood-rearing was among the longest reported for any diurnal raptor. Twenty-eight nests were found in a variety of native and exotic habitats. Incubation lasted 38 days, nestlingsfledged after 59-63 days, and parentscared for fledglings for an average of 30.2 weeks, which was 2.5 to 10 times longer than similar-size temperate zone raptors. Males assisted females with incubation, but only females brooded young. Radio-tagged juveniles remained within 0.63 km of their nests for the first two months after fledging, after which dispersal distances expanded gradually. Avian (45%) and mammalian (54%) prey dominated the diet of nestlings. There was no evidence that avian malaria, introduced predators, or environmental contaminantswere affecting their population. Based upon es- timates of population size, the availability of suitable nesting habitat, and reproductivesuc- cess, we suggestthe speciesbe consideredfor downlisting from endangeredto threatened status. Key words: breeding ecology,Buteo solitarius, clutch size, diet, Hawaiian Hawk, nest success. -

Alpha Codes for 2168 Bird Species (And 113 Non-Species Taxa) in Accordance with the 62Nd AOU Supplement (2021), Sorted Taxonomically
Four-letter (English Name) and Six-letter (Scientific Name) Alpha Codes for 2168 Bird Species (and 113 Non-Species Taxa) in accordance with the 62nd AOU Supplement (2021), sorted taxonomically Prepared by Peter Pyle and David F. DeSante The Institute for Bird Populations www.birdpop.org ENGLISH NAME 4-LETTER CODE SCIENTIFIC NAME 6-LETTER CODE Highland Tinamou HITI Nothocercus bonapartei NOTBON Great Tinamou GRTI Tinamus major TINMAJ Little Tinamou LITI Crypturellus soui CRYSOU Thicket Tinamou THTI Crypturellus cinnamomeus CRYCIN Slaty-breasted Tinamou SBTI Crypturellus boucardi CRYBOU Choco Tinamou CHTI Crypturellus kerriae CRYKER White-faced Whistling-Duck WFWD Dendrocygna viduata DENVID Black-bellied Whistling-Duck BBWD Dendrocygna autumnalis DENAUT West Indian Whistling-Duck WIWD Dendrocygna arborea DENARB Fulvous Whistling-Duck FUWD Dendrocygna bicolor DENBIC Emperor Goose EMGO Anser canagicus ANSCAN Snow Goose SNGO Anser caerulescens ANSCAE + Lesser Snow Goose White-morph LSGW Anser caerulescens caerulescens ANSCCA + Lesser Snow Goose Intermediate-morph LSGI Anser caerulescens caerulescens ANSCCA + Lesser Snow Goose Blue-morph LSGB Anser caerulescens caerulescens ANSCCA + Greater Snow Goose White-morph GSGW Anser caerulescens atlantica ANSCAT + Greater Snow Goose Intermediate-morph GSGI Anser caerulescens atlantica ANSCAT + Greater Snow Goose Blue-morph GSGB Anser caerulescens atlantica ANSCAT + Snow X Ross's Goose Hybrid SRGH Anser caerulescens x rossii ANSCAR + Snow/Ross's Goose SRGO Anser caerulescens/rossii ANSCRO Ross's Goose -

Repeated Sequence Homogenization Between the Control and Pseudo
Repeated Sequence Homogenization Between the Control and Pseudo-Control Regions in the Mitochondrial Genomes of the Subfamily Aquilinae LUIS CADAH´IA1*, WILHELM PINSKER1, JUAN JOSE´ NEGRO2, 3 4 1 MIHAELA PAVLICEV , VICENTE URIOS , AND ELISABETH HARING 1Molecular Systematics, 1st Zoological Department, Museum of Natural History Vienna, Vienna, Austria 2Department of Evolutionary Ecology, Estacio´n Biolo´gica de Don˜ana, Sevilla, Spain 3Centre of Ecological and Evolutionary Synthesis (CEES), Department of Biology, Faculty for Natural Sciences and Math, University of Oslo, Oslo, Norway 4Estacio´n Biolo´gica Terra Natura (Fundacio´n Terra Natura—CIBIO), Universidad de Alicante, Alicante, Spain ABSTRACT In birds, the noncoding control region (CR) and its flanking genes are the only parts of the mitochondrial (mt) genome that have been modified by intragenomic rearrangements. In raptors, two noncoding regions are present: the CR has shifted to a new position with respect to the ‘‘ancestral avian gene order,’’ whereas the pseudo-control region (CCR) is located at the original genomic position of the CR. As possible mechanisms for this rearrangement, duplication and transposition have been considered. During characterization of the mt gene order in Bonelli’s eagle Hieraaetus fasciatus, we detected intragenomic sequence similarity between the two regions supporting the duplication hypothesis. We performed intra- and intergenomic sequence comparisons in H. fasciatus and other falconiform species to trace the evolution of the noncoding mtDNA regions in Falconiformes. We identified sections displaying different levels of similarity between the CR and CCR. On the basis of phylogenetic analyses, we outline an evolutionary scenario of the underlying mutation events involving duplication and homogenization processes followed by sporadic deletions. -

Diurnal Birds of Prey of Belize
DIURNAL BIRDS OF PREY OF BELIZE Nevertheless, we located thirty-four active Osprey by Dora Weyer nests, all with eggs or young. The average number was three per nest. Henry Pelzl, who spent the month The Accipitridae of June, 1968, studying birds on the cayes, estimated 75 Belize is a small country south of the Yucatán to 100 pairs offshore. Again, he could not get to many Peninsula on the Caribbean Sea. Despite its small of the outer cayes. It has been reported that the size, 285 km long and 112 km wide (22 963 km2), southernmost part of Osprey range here is at Belize encompasses a great variety of habitats: Dangriga (formerly named Stann Creek Town), a mangrove cays and coastal forests, lowland tropical little more than halfway down the coast. On Mr pine/oak/palm savannas (unique to Belize, Honduras Knoder’s flight we found Osprey nesting out from and Nicaragua), extensive inland marsh, swamp and Punta Gorda, well to the south. lagoon systems, subtropical pine forests, hardwood Osprey also nest along some of the rivers inland. Dr forests ranging from subtropical dry to tropical wet, Stephen M. Russell, author of A Distributional Study and small areas of elfin forest at the top of the highest of the Birds of British Honduras, the only localized peaks of the Maya Mountains. These mountains are reference, in 1963, suspects that most of the birds seen built of extremely old granite overlaid with karst inland are of the northern race, carolinensis, which limestone. The highest is just under 1220 m. Rainfall winters here.