SYT-SSX1 and SYT-SSX2 Interfere with Repression of E-Cadherin by Snail and Slug
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

SS18 (SYT) (18Q11) Gene Rearrangement by FISH Indications for Ordering Genetics
SS18 (SYT) (18q11) Gene Rearrangement by FISH Indications for Ordering Genetics Diagnosis of synovial sarcoma in conjunction with histologic Translocations – SS18-SSX1, SS18-SSX2 and clinical information Structure/function Test Description • SS18 is located on chromosome 18 • SSX1 and SSX2 are located on the X-chromosome Fluorescence in situ hybridization • Each gene in the translocation codes for proteins that have opposite transcriptional functions Tests to Consider o SS18 – activator of oncogenes Primary test o SSX1, SSX2 – tumor suppression SS18 (SYT) (18q11) Gene Rearrangement by FISH 3001303 Test Interpretation • Molecular diagnosis of synovial sarcoma Results Related test • Positive – SS18 rearrangement is detected Chromosome FISH, Interphase 2002298 o SSX translocation partner is not identified with this • Specific probe for SS18 (SYT) rearrangement must be testing methodology requested o Synovial sarcoma likely • Fresh tissue specimens only • Negative – no SS18 rearrangement detected Disease Overview o Does not entirely exclude the presence of an SS18 rearrangement as some translocations are cryptic and Incidence – rare not evaluable by this testing methodology • Synovial sarcomas account for 8-10% of all soft tissue o Does not entirely exclude diagnosis of synovial sarcoma sarcomas Limitations Diagnostic/prognostic issues • Testing using tissue fixed in alcohol-based or non-formalin • Synovial sarcomas may resemble other neoplasms, fixatives has not been validated using this method particularly those displaying an epithelioid, spindle cell, or • SS18 fusion partners are not detected with this test combined morphology • t(X;18)(p11.2;q11.2) translocation serves as a specific marker for synovial sarcoma o SS18 (SYT) gene fuses with SSX gene . Fusion with SSX1 in ~65% of synovial sarcomas . -

A Novel Type of SYT/SSX Fusion
A Novel Type of SYT/SSX Fusion: Methodological and Biological Implications Maria Törnkvist, M.Sc., Bertha Brodin, Ph.D., Armando Bartolazzi, M.D., Ph.D., Olle Larsson, M.D., Ph.D. Department of Cellular and Molecular Tumor Pathology, Cancer Centrum Karolinska, Karolinska Hospital (MT, BB, AB, OL), Stockholm, Sweden; and Department of Pathology, Regina Elena Cancer Institute (AB), Rome, Italy the presence of SYT/SSX transcripts in two cases Synovial sarcoma (SS) is a rare soft-tissue tumor using the proposed RT-PCR approach. Applications that affects children and young adults. It is charac- of optimized RT-PCR can contribute to reduce false- terized by the chromosomal translocation t(X; negative SYT/SSX SS cases reported in literature. 18)(p11.2;q11.2), which results in the fusion of the SYT gene on chromosome 18 with a SSX gene on KEY WORDS: RT-PCR, Synovial sarcoma, SYT/SSX chromosome X. In the majority of cases, SYT is fusion gene, SYT/SSX variants. fused to exon 5 of SSX1 (64%), SSX2 (36%), or, Mod Pathol 2002;15(6):679–685 rarely, SSX4. A novel fusion transcript variant deriv- ing from the fusion of SYT to exon 6 of SSX4 gene Synovial sarcomas (SS) account for 7 to 10% of all (SYT/SSX4v) was found coexpressed in one of the human soft-tissue sarcomas and are mainly located previously reported SYT/SSX4 cases. In the present in the extremities in the vicinity of large joints (1). investigation, we describe a new SS case that was Depending on histomorphological appearance, SSs previously shown to be negative for SYT/SSX1 and are usually subdivided into two major forms, bipha- SYT/SSX2 expression by conventional reverse tran- sic and monophasic. -

A Novel FISH Assay for SS18–SSX Fusion Type in Synovial Sarcoma
Laboratory Investigation (2004) 84, 1185–1192 & 2004 USCAP, Inc All rights reserved 0023-6837/04 $30.00 www.laboratoryinvestigation.org A novel FISH assay for SS18–SSX fusion type in synovial sarcoma Cecilia Surace1,2, Ioannis Panagopoulos1, Eva Pa˚lsson1, Mariano Rocchi2, Nils Mandahl1 and Fredrik Mertens1 1Department of Clinical Genetics, Lund University Hospital, Lund, Sweden and 2DAPEG, Section of Genetics, University of Bari, Bari, Italy Synovial sarcoma is a morphologically, clinically and genetically distinct entity that accounts for 5–10% of all soft tissue sarcomas. The t(X;18)(p11.2;q11.2) is the cytogenetic hallmark of synovial sarcoma and is present in more than 90% of the cases. It produces three types of fusion gene formed in part by SS18 from chromosome 18 and by SSX1, SSX2 or, rarely, SSX4 from the X chromosome. The SS18–SSX fusions do not seem to occur in other tumor types, and it has been shown that in synovial sarcoma a clear correlation exists between the type of fusion gene and histologic subtype and, more importantly, clinical outcome. Previous analyses regarding the type of fusion genes have been based on PCR amplification of the fusion transcript, requiring access to good- quality RNA. In order to obtain an alternative tool to diagnose and follow this malignancy, we developed a fluorescence in situ hybridization (FISH) assay that could distinguish between the two most common fusion genes, that is, SS18–SSX1 and SS18–SSX2. The specificity of the selected bacterial artificial chromosome clones used in the detection of these fusion genes, as well as the sensitivity of the analysis in metaphase and interphase cells, was examined in a series of 28 synovial sarcoma samples with known fusion gene status. -

Gene Section Review
Atlas of Genetics and Cytogenetics in Oncology and Haematology OPEN ACCESS JOURNAL AT INIST-CNRS Gene Section Review SSX2 (Synovial Sarcoma, X breakpoint 2) Josiane Eid, Christina Garcia, Andrea Frump Department of Cancer Biology, Vanderbilt University Medical Center, Nashville, TN 37232, USA (JE, CG, AF) Published in Atlas Database: April 2008 Online updated version: http://AtlasGeneticsOncology.org/Genes/SSX2ID42406chXp11.html DOI: 10.4267/2042/44431 This work is licensed under a Creative Commons Attribution-Noncommercial-No Derivative Works 2.0 France Licence. © 2009 Atlas of Genetics and Cytogenetics in Oncology and Haematology detected in liver, testis, skin melanoma, endometrium, Identity choriocarcinoma, placenta, spleen of Hodgkins Other names: CT5.2; HD21; HOM-MEL-40; lymphoma. MGC119055; MGC15364; MGC3884; RP11-552J9.2; SSX; SSX2A; SSX2B Protein HGNC (Hugo): SSX2 Location: Xp11.22 Description So far, two SSX2 protein isoforms (a and b) are known DNA/RNA to exist. Their mRNAs correspond to SV1 (1466 bases) and SV3 (1322 bases) splice variants, respectively. The Description start codon for both isoforms is located in Exon 2. The SSX2 gene locus encompasses 9 exons and 10,304 SSX2 isoform a is 233 amino acids (26.5 kD) and bp (Xp11; 52,752,974-52,742,671). SSX2 isoform b 188 amino acids (21.6 kD). Of both isoforms, SSX2 isoform b is the most commonly seen Transcription and so far the best studied. The SSX2 gene is transcribed on the minus strand. 7 SSX2 mRNA splice variants (SV1-SV7) have been SSX2 Locus and mRNA Splice Variants. Note: Exons are drawn to scale. Atlas Genet Cytogenet Oncol Haematol. -

SSX2 Is Differentially Expressed in Models of MERS Coronavirus-PDF 042820
1 SSX2 is differentially expressed in models of MERS coronavirus infection. 2 Shahan Mamoor, MS1 1Thomas Jefferson School of Law 3 San Diego, CA 92101 4 [email protected] 5 The coronavirus COVID19 pandemic is an emerging biosafety threat to the nation and the 6 world (1). There are no treatments approved for coronavirus infection in humans (2) and there is a lack of information available regarding the basic transcriptional behavior of human cells 7 and mammalian tissues following coronavirus infection. We mined multiple independent public 8 (3) or published datasets (4-8) containing transcriptome data from infection models of human coronavirus 229E, the severe acute respiratory syndrome (SARS) coronavirus and Middle East 9 respiratory syndrome (MERS) coronavirus to discover genes whose differential expression was conserved across the coronavirus family. We identified SSX2 (9) as a differentially expressed 10 gene following infection of human cells specifically with two types of MERS coronaviruses. and not after infection of human cells with human coronavirus 229E, or and in the lungs of mice 11 and ferrets infected with SARS coronavirus. An SSX2 interacting protein, SSX2IP, was among the genes most differentially expressed in the ferret blood after infection with SARS 12 coronavirus. The expression of SSX2 is modulated to a degree unlike most any other gene 13 following infection with MERS coronaviruses. 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Keywords: SSX2, coronavirus, MERS coronavirus, SARS coronavirus, human coronavirus 28 229E, SARS-CoV-2, COVID19, systems biology of viral infection. 1 1 Viruses are classified according to a system known as the “Baltimore” classification of 2 viruses (10) wherein the characteristics of the viral genome - whether it is positive-sense or 3 negative-sense, whether it is single-stranded or double-stranded, whether it is composed or 4 RNA or DNA - are used to group viruses into families. -

Product Sheet CA1235
SSX2 Antibody Applications: WB, IHC Detected MW: 25 kDa Cat. No. CA1235 Species & Reactivity: Human, Mouse, Rat Isotype: Rabbit IgG BACKGROUND SSX2 belongs to the family of highly homologous based immunotherapy.4 Two transcript variants synovial sarcoma X (SSX) breakpoint proteins. encoding distinct isoforms have been identified for The SSX gene family is composed of at least 9 SSX2 gene. SSX2 is thought to function in functional and highly homologous members and development and germ line cells as a repressive shown to be located on chromosome X. The gene regulator. Its control of gene expression is normal testis expresses SSX1, 2, 3, 4, 5, and 7, believed to be epigenetic in nature and to involve but not 6, 8, or 9. In tumors, SSX1, 2, and 4 are chromatin modification and remodeling. It is most expressed at varying frequencies, whereas SSX3, likely mediated by the association of SSX2 with the 5, and 6 are rarely expressed. In addition, no Polycomb gene silencing complex at the SSXRD expression of SSX8, or 9 has been observed. SSX1 domain. Polycomb silencing involves chromatin to SSX5 are also normally expressed in thyroid.1 compaction, DNA methylation, repressive histone The SSX family shares nucleotide homology modifications and inaccessibility of promoter ranging from 88% to 95%, and amino acid regions to transcription machineries. Other SSX2- homology ranging from 77% to 91%. The NH2- interacting partners include the LIM homeobox terminal moieties of the SSX proteins exhibit protein LHX4, a Ras-like GTPase Interactor, homology to the Krüppel-associated box (KRAB) RAB3IP thought to be involved in vesicular domain, a domain that is known to be involved in transport, and SSX2IP, a putative cell transcriptional repression. -

Identification of SYT-SSX Transcripts from Synovial Sarcomas Using RT-Multiplex PCR and Capillary Electrophoresis
Modern Pathology (2006) 19, 641–647 & 2006 USCAP, Inc All rights reserved 0893-3952/06 $30.00 www.modernpathology.org Identification of SYT-SSX transcripts from synovial sarcomas using RT-multiplex PCR and capillary electrophoresis John A Thorson, Helmut C Weigelin, Robert E Ruiz, Jennifer K Howard and David R Lucas Department of Pathology, University of Michigan, Ann Arbor, MI, USA Synovial sarcomas are highly malignant tumors of soft tissue which are characterized by the t(X;18) resulting in SYT-SSX fusion transcript production. Diagnosis of these tumors based on histology can be challenging, particularly when minimal biopsy specimens are presented to the pathologist. Demonstration by molecular methods of SYT-SSX transcripts is a useful adjunct for diagnosis in these situations. We have developed an assay, which combines one-step RT-multiplex PCR with capillary electrophoresis to detect and genotype the SYT-SSX transcripts from synovial sarcomas. Small amplicons from chimeric transcripts as well as GAPD transcripts are differentially labeled with fluorophores, allowing detection and size discrimination by capillary electrophoresis. In a study of 32 formalin-fixed soft tissue tumor specimens, the assay detected chimeric transcripts from 17/22 (77%) synovial sarcomas. All five assay negative specimens yielded no intact RNA as evidenced by lack of a GAPD amplicon. Chimeric transcripts were not detected in 9/9 malignant peripheral nerve sheath tumors or 1/1 epithelioid sarcoma. Representative amplicons were sequenced and confirmed the genotype results obtained by capillary electrophoresis. One-step RT-multiplex PCR combined with capillary electrophoresis is a rapid and accurate method for the detection and genotypic classification of SYT-SSX transcripts from fixed tissue specimens. -

Genomic and Expression Profiling of Human Spermatocytic Seminomas: Primary Spermatocyte As Tumorigenic Precursor and DMRT1 As Candidate Chromosome 9 Gene
Research Article Genomic and Expression Profiling of Human Spermatocytic Seminomas: Primary Spermatocyte as Tumorigenic Precursor and DMRT1 as Candidate Chromosome 9 Gene Leendert H.J. Looijenga,1 Remko Hersmus,1 Ad J.M. Gillis,1 Rolph Pfundt,4 Hans J. Stoop,1 Ruud J.H.L.M. van Gurp,1 Joris Veltman,1 H. Berna Beverloo,2 Ellen van Drunen,2 Ad Geurts van Kessel,4 Renee Reijo Pera,5 Dominik T. Schneider,6 Brenda Summersgill,7 Janet Shipley,7 Alan McIntyre,7 Peter van der Spek,3 Eric Schoenmakers,4 and J. Wolter Oosterhuis1 1Department of Pathology, Josephine Nefkens Institute; Departments of 2Clinical Genetics and 3Bioinformatics, Erasmus Medical Center/ University Medical Center, Rotterdam, the Netherlands; 4Department of Human Genetics, Radboud University Medical Center, Nijmegen, the Netherlands; 5Howard Hughes Medical Institute, Whitehead Institute and Department of Biology, Massachusetts Institute of Technology, Cambridge, Massachusetts; 6Clinic of Paediatric Oncology, Haematology and Immunology, Heinrich-Heine University, Du¨sseldorf, Germany; 7Molecular Cytogenetics, Section of Molecular Carcinogenesis, The Institute of Cancer Research, Sutton, Surrey, United Kingdom Abstract histochemistry, DMRT1 (a male-specific transcriptional regulator) was identified as a likely candidate gene for Spermatocytic seminomas are solid tumors found solely in the involvement in the development of spermatocytic seminomas. testis of predominantly elderly individuals. We investigated these tumors using a genome-wide analysis for structural and (Cancer Res 2006; 66(1): 290-302) numerical chromosomal changes through conventional kar- yotyping, spectral karyotyping, and array comparative Introduction genomic hybridization using a 32 K genomic tiling-path Spermatocytic seminomas are benign testicular tumors that resolution BAC platform (confirmed by in situ hybridization). -
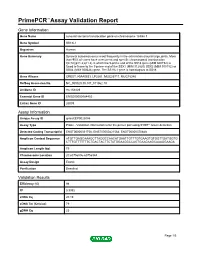
Primepcr™Assay Validation Report
PrimePCR™Assay Validation Report Gene Information Gene Name synovial sarcoma translocation gene on chromosome 18-like 1 Gene Symbol SS18L1 Organism Human Gene Summary Synovial sarcomas occur most frequently in the extremities around large joints. More than 90% of cases have a recurrent and specific chromosomal translocation t(X;18)(p11.2;q11.2) in which the 5-prime end of the SS18 gene (MIM 600192) is fused in-frame to the 3-prime end of the SSX1 (MIM 312820) SSX2 (MIM 300192) or SSX4 (MIM 300326) gene. The SS18L1 gene is homologous to SS18. Gene Aliases CREST, KIAA0693, LP2261, MGC26711, MGC78386 RefSeq Accession No. NC_000020.10, NT_011362.10 UniGene ID Hs.154429 Ensembl Gene ID ENSG00000184402 Entrez Gene ID 26039 Assay Information Unique Assay ID qHsaCEP0025086 Assay Type Probe - Validation information is for the primer pair using SYBR® Green detection Detected Coding Transcript(s) ENST00000331758, ENST00000421564, ENST00000370848 Amplicon Context Sequence ATGTTGAGCAAAGCTTAGGCCAACATGAATTGTTTGTGAAGTGTGGTTGATGGTG CTTTGTTTTTTTCTGACTACTTCTATGGAAGGCCAGTGAAGAAGCAAAGGAAGA Amplicon Length (bp) 79 Chromosome Location 20:60756836-60756944 Assay Design Exonic Purification Desalted Validation Results Efficiency (%) 98 R2 0.9992 cDNA Cq 20.19 cDNA Tm (Celsius) 79 gDNA Cq 25 Page 1/5 PrimePCR™Assay Validation Report Specificity (%) 100 Information to assist with data interpretation is provided at the end of this report. Page 2/5 PrimePCR™Assay Validation Report SS18L1, Human Amplification Plot Amplification of cDNA generated from 25 ng of universal -

Thesis for Word XP
Thesis for doctoral degree (Ph.D.) 2008 Thesis for doctoral degree (Ph.D.) 2008 Molecular Mechanisms Underlying the Oncogenic Function of SS18 and SSX Molecular Mechanisms Underlying the Oncogenic Function of SS18 and SSX Pádraig D’Arcy Fredrik Bredin Pádraig D’Arcy Department of Oncology-Pathology Cancer Center Karolinska Karolinska Institutet, Stockholm, Sweden MOLECULAR MECHANISMS UNDERLYING THE ONCOGENIC FUNCTION OF SS18 AND SSX. Pádraig D'Arcy Stockholm 2008 All previously published papers were reproduced with permission from the publisher. Published by Karolinska Institutet. Printed by Larserics Digital Print AB © Pádraig D'Arcy, 2008 ISBN 978-91-7357-481-5 I’m digging for fire The Pixies To my parents Abstract The SS18 and SSX genes were initially identified based on their reoccurrence as fusion partners in synovial sarcoma. As a result of the specific chromosomal translocation t(X:18), the SS18 gene from chromosome 18 becomes fused with members of the SSX gene family on the X chromosome resulting in the generation of a novel chimeric fusion gene SS18-SSX. The SS18 gene encodes a ubiquitously expressed transcriptional activator, whereas the SSX gene encodes a transcriptional repressor whose expression is restricted to germ cells and numerous cancers. Thus, the resultant SS18-SSX fusion gene encodes a transcription factor with dual trans activation and repression properties; the expression of which is the initiating event of synovial sarcoma. We present the findings that SSX, along with several other members of the CT-antigen family is expressed in mesenchymal stem cells and their expression is down regulated following differentiation. Knockdown of SSX could effectively impair cell migration, a phenotype associated with down regulation of MMP2 expression adding a functional role for SSX in stem and tumor cell migration. -

The Impact of Chromosomal Translocation Locus and Fusion Oncogene Coding Sequence in Synovial Sarcomagenesis
HHS Public Access Author manuscript Author ManuscriptAuthor Manuscript Author Oncogene Manuscript Author . Author manuscript; Manuscript Author available in PMC 2016 September 23. Published in final edited form as: Oncogene. 2016 September 22; 35(38): 5021–5032. doi:10.1038/onc.2016.38. The impact of chromosomal translocation locus and fusion oncogene coding sequence in synovial sarcomagenesis Kevin B. Jones1,2,3,*, Jared J. Barrott1,2,3, Mingchao Xie4, Malay Haldar5, Huifeng Jin1,2,3, Ju-Fen Zhu1,2,3, Michael J. Monument1,3, Tim L. Mosbruger3,6, Ellen M. Langer5, R. Lor Randall1,3, Richard K. Wilson4,7,8,9, Bradley R. Cairns2,3,10, Li Ding4,7,8,9, and Mario R. Capecchi5 1Department of Orthopaedics, University of Utah, Salt Lake City, Utah 84112, USA 2Department of Oncological Sciences, University of Utah, Salt Lake City, Utah 84112, USA 3Huntsman Cancer Institute, University of Utah, Salt Lake City, Utah 84112, USA 4Department of Medicine, Washington University, St. Louis, Missouri 63108, USA 5Department of Human Genetics, University of Utah, Salt Lake City, Utah, USA 6Deparment of Bioinformatics, University of Utah, Salt Lake City, Utah, 84112, USA 7McDonnell Genome Institute, Washington University, St. Louis, Missouri 63108, USA 8Department of Genetics, Washington University, St. Louis, Missouri 63108, USA 9Siteman Cancer Center, Washington University, St. Louis, Missouri 63108, USA 10Howard Hughes Medical Institute, University of Utah, Salt Lake City, UT 84112, USA Abstract Synovial sarcomas are aggressive soft-tissue malignancies that express chromosomal translocation-generated fusion genes, SS18-SSX1 or SS18-SSX2 in most cases. Here, we report a mouse sarcoma model expressing SS18-SSX1, complementing our prior model expressing SS18- SSX2. -

Fusions of the SYT and SSX Genes in Synovial Sarcoma
Oncogene (2001) 20, 5755 ± 5762 ã 2001 Nature Publishing Group All rights reserved 0950 ± 9232/01 $15.00 www.nature.com/onc Fusions of the SYT and SSX genes in synovial sarcoma Marc Ladanyi*,1 1Department of Pathology, Memorial Sloan-Kettering Cancer Center, New York, NY, USA Synovial sarcomas are high grade spindle cell tumors tumors. Ultrastructurally, the epithelial cells lining the that are divided into two major histologic subtypes, glandlike spaces in biphasic tumors show luminal biphasic and monophasic, according to the respective microvilli and are interconnected by various types of presence or absence of a well-developed glandular cell junctions. epithelial component. They contain in essentially all cases a t(X;18) representing the fusion of SYT (at 18q11) with either SSX1 or SSX2 (both at Xp11). Fusions of SYT with SSX1 or SSX2 in synovial Neither SYT, nor the SSX proteins contain DNA- sarcoma: prevalence and structural aspects binding domains. Instead, they appear to be transcrip- tional regulators whose actions are mediated primarily A t(X;18) is detected cytogenetically in over 90% of through protein-protein interactions, with BRM in the synovial sarcomas, regardless of histologic subtype. In case of SYT, and with Polycomb group repressors in the relation to this recurrent X chromosome translocation, case of SSX. Ongoing work on the SYT ± SSX fusion it is of incidental interest to note that the male to and synovial sarcoma should yield a variety of data of female ratio for this tumor is approximately even. broader biological interest, in areas such as BRM and Cloning of the translocation breakpoints initially Polycomb group function and dysfunction, transcrip- showed that the t(X;18) results in the fusion of two tional targets of SYT ± SSX proteins and their native novel genes, designated SYT (at 18q11) and SSX (at counterparts, dierential gene regulation by SYT ± SSX1 Xp11) (Figure 1) (Clark et al., 1994).