Microfossils in Metasediments from Prins Karls Forland, Western Svalbard
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Handbok07.Pdf
- . - - - . -. � ..;/, AGE MILL.YEAR$ ;YE basalt �- OUATERNARY votcanoes CENOZOIC \....t TERTIARY ·· basalt/// 65 CRETACEOUS -� 145 MESOZOIC JURASSIC " 210 � TRIAS SIC 245 " PERMIAN 290 CARBONIFEROUS /I/ Å 360 \....t DEVONIAN � PALEOZOIC � 410 SILURIAN 440 /I/ ranite � ORDOVICIAN T 510 z CAM BRIAN � w :::;: 570 w UPPER (J) PROTEROZOIC � c( " 1000 Ill /// PRECAMBRIAN MIDDLE AND LOWER PROTEROZOIC I /// 2500 ARCHEAN /(/folding \....tfaulting x metamorphism '- subduction POLARHÅNDBOK NO. 7 AUDUN HJELLE GEOLOGY.OF SVALBARD OSLO 1993 Photographs contributed by the following: Dallmann, Winfried: Figs. 12, 21, 24, 25, 31, 33, 35, 48 Heintz, Natascha: Figs. 15, 59 Hisdal, Vidar: Figs. 40, 42, 47, 49 Hjelle, Audun: Figs. 3, 10, 11, 18 , 23, 28, 29, 30, 32, 36, 43, 45, 46, 50, 51, 52, 53, 54, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 71, 72, 75 Larsen, Geir B.: Fig. 70 Lytskjold, Bjørn: Fig. 38 Nøttvedt, Arvid: Fig. 34 Paleontologisk Museum, Oslo: Figs. 5, 9 Salvigsen, Otto: Figs. 13, 59 Skogen, Erik: Fig. 39 Store Norske Spitsbergen Kulkompani (SNSK): Fig. 26 © Norsk Polarinstitutt, Middelthuns gate 29, 0301 Oslo English translation: Richard Binns Editor of text and illustrations: Annemor Brekke Graphic design: Vidar Grimshei Omslagsfoto: Erik Skogen Graphic production: Grimshei Grafiske, Lørenskog ISBN 82-7666-057-6 Printed September 1993 CONTENTS PREFACE ............................................6 The Kongsfjorden area ....... ..........97 Smeerenburgfjorden - Magdalene- INTRODUCTION ..... .. .... ....... ........ ....6 fjorden - Liefdefjorden................ 109 Woodfjorden - Bockfjorden........ 116 THE GEOLOGICAL EXPLORATION OF SVALBARD .... ........... ....... .......... ..9 NORTHEASTERN SPITSBERGEN AND NORDAUSTLANDET ........... 123 SVALBARD, PART OF THE Ny Friesland and Olav V Land .. .123 NORTHERN POLAR REGION ...... ... 11 Nordaustlandet and the neigh- bouring islands........................... 126 WHA T TOOK PLACE IN SVALBARD - WHEN? .... -

Foraminifera1 Stratigraphy of Raised Marine Deposits, Representing Isotope Stage 5, Prins Karls Forland, Western Sval Bard
Foraminifera1 stratigraphy of raised marine deposits, representing isotope stage 5, Prins Karls Forland, western SVal bard HELENE BERGSTEN. TORBJORN ANDERSON and OLAFUR INGOLFSSON Bergsten, H., Andersson. T. & Ingolfsson, 0. 1998: Foraminiferd stratigraphy of raised marine deposits, representing isotope stage 5. Prins Karls Forland, western Svalbard. Polar Research 17(1). 81-91. Two raised marine sequences from Prim Karls Forland, western Svalbard, interpreted to have been deposited during part of isotope substage Se (Eemian) and substage 5a, were studied for foraminifera content. Time constraints are given by I4C ages, infrared stimulated luminescence age estimates and amino acid ratios in suhfossil marine molluscs. A diamicton (unit B) separates the two marine sequences and reflects an advancement of local glaciers sometime late in isotope stage 5. The two marine sequences contain diverse benthic foraminifera1 faunas, indicating periods of a relatively warm and seasonally ice-free marine shelf environment. Compared to the lowermost sequence (unit A), the upper marine sequence (unit C) seems to reflect a more shallow environment that could have resulted from the global lowering of the sea level towards the end of isotope stage 5. Our results further emphasise the problem of biostratigraphic distinction between interglacial and interstadial deposits at high latitudes, with temperature conditions for substage Sa close to those of substage 5e and present conditions. Helene Bergsten, Torbjorn Andersson and Olufur Ingdlfsson, Earth Sciences Centre, Goteborg University, Box 460, S-405 30 Goteborg, Sweden. Introduction Chronology and glacial history The Poolepynten site is situated on Prins Karls Sediments in the Poolepynten sections are ex- Forland, western Svalbard at N78'27'El l"40' posed in up to 10 metre high coastal cliffs over a (Fig. -

ESG Perspectives June 2019 Svalbard Sojourn an Arctic
June 2019 ESG Perspectives ™ SVALBARD SOJOURN: AN ARCTIC EXPERIENCE by Bob Smith, President & CIO islands are 60% glaciated with some of the world’s fastest-moving glaciers. The balance of the region is 30% barren ground, and 10% is covered with very low ground vegetation. In contrast to other Arctic regions, the Svalbard has no indigenous population and there is no historical evidence that the Vikings settled in the area during their time. In fact, it was the 1596 Dutch expedition of Willem Barents who discovered and drew maps of the region before his ship was crushed by freezing sea ice leading to his untimely death. However, the work of this expedition survived, and it led to the exploration of the region by other European countries over the Photo of Bob Smith in Kungsfjord, Svalbard centuries. This eventually gave rise to the exploitation It is said that what happens in the Arctic doesn’t stay of the natural resources of the region as a destination in the Arctic. That is because this region provides for whalers, fur trappers, and seal hunters, as well as essential global climate regulation and substantial other animal-based products. With the industrialization ecosystem benefits to humanity outside and beyond of Europe and the arrival of steel-hulled ships in the its boundaries. Indeed, the Arctic environment, and early 1900s, this region also eventually became a source human society and its economic activities are deeply for industrial minerals and, in particular, coal. connected to each other, representing a pivotal link in a complex adaptive global ecosystem. -

Chapter 1 Svalbard, 3 Chapter 2 Outline History of Geological Research, 16 Chapter 3 Svalbard's Geological Frame, 23
Downloaded from http://mem.lyellcollection.org/ by guest on September 25, 2021 PART 1 Introduction Chapter 1 Svalbard, 3 Chapter 2 Outline history of geological research, 16 Chapter 3 Svalbard's geological frame, 23 Mid-season view of the glaciers Comfortlessbreen (on the left) and Aavartsmarkbreen (beyond). The rocks are Early Vendian with Varanger tillites. Stratigraphic sections are generally worked along the glacier margins either by porterage from the shore (in this case Egelskbukta) or by sledge from the interior. Photo M. J. Hambrey (SP. 455). Late season view from the east over the terminal crevassed glacier Monacobreen. In this case access up the glacier is almost impossible because the lower reaches are deeply crevassed and the glacier terminates in ice-cliffs in inner Liefdefjorden. The glacier beyond offers an easy route westward. Photo P. W. Webb, CSE 1989. Downloaded from http://mem.lyellcollection.org/ by guest on September 25, 2021 View from Ossian Sarsfjellet at the eastern end of Kongsfjorden towards the mountains of Broggerhalvoya which are reflected in the fjord. The intervening fjord carries a scatter of small bergs which have calved from the glacier cliffs of Kronebreen and Kongsbreen respectively to north and south of the photographer. The concentration of ice depends on wind and tide and is navigable with care in a slow moving boat. The small bergs melt rapidly in the summer. Photo M. J. Hambrey (SP96.122) 1996. The CSE motorboat Salterella in mid-Kongsfjorden seen when looking out to sea with the landmark Kapp Mitra to the right where the rocks are Caledonian metamorphosed basement of pre-Vendian rocks. -

Protected Areas in Svalbard – Securing Internationally Valuable Cultural and Natural Heritage Contents Preface
Protected areas in Svalbard – securing internationally valuable cultural and natural heritage Contents Preface ........................................................................ 1 – Moffen Nature Reserve ......................................... 13 From no-man’s-land to a treaty and the Svalbard – Nordaust-Svalbard Nature Reserve ...................... 14 Environmental Protection Act .................................. 4 – Søraust-Svalbard Nature Reserve ......................... 16 The history of nature and cultural heritage – Forlandet National Park .........................................18 protection in Svalbard ................................................ 5 – Indre Wijdefjorden National Park ......................... 20 The purpose of the protected areas .......................... 6 – Nordenskiöld Land National Park ........................ 22 Protection values ........................................................ 7 – Nordre Isfjorden National Park ............................ 24 Nature protection areas in Svalbard ........................10 – Nordvest-Spitsbergen National Park ................... 26 – Bird sanctuaries ..................................................... 11 – Sassen-Bünsow Land National Park .................... 28 – Bjørnøya Nature Reserve ...................................... 12 – Sør-Spitsbergen National Park ..............................30 – Ossian Sars Nature Reserve ................................. 12 Svalbard in a global context ..................................... 32 – Hopen Nature Reserve -

Svalbard 2015–2016 Meld
Norwegian Ministry of Justice and Public Security Published by: Norwegian Ministry of Justice and Public Security Public institutions may order additional copies from: Norwegian Government Security and Service Organisation E-mail: [email protected] Internet: www.publikasjoner.dep.no KET T Meld. St. 32 (2015–2016) Report to the Storting (white paper) Telephone: + 47 222 40 000 ER RY M K Ø K J E L R I I Photo: Longyearbyen, Tommy Dahl Markussen M 0 Print: 07 PrintMedia AS 7 9 7 P 3 R 0 I 1 08/2017 – Impression 1000 N 4 TM 0 EDIA – 2 Svalbard 2015–2016 Meld. St. 32 (2015–2016) Report to the Storting (white paper) 1 Svalbard Meld. St. 32 (2015–2016) Report to the Storting (white paper) Svalbard Translation from Norwegian. For information only. Table of Contents 1 Summary ........................................ 5 6Longyearbyen .............................. 39 1.1 A predictable Svalbard policy ........ 5 6.1 Introduction .................................... 39 1.2 Contents of each chapter ............... 6 6.2 Areas for further development ..... 40 1.3 Full overview of measures ............. 8 6.2.1 Tourism: Longyearbyen and surrounding areas .......................... 41 2Background .................................. 11 6.2.2 Relocation of public-sector jobs .... 43 2.1 Introduction .................................... 11 6.2.3 Port development ........................... 44 2.2 Main policy objectives for Svalbard 11 6.2.4 Svalbard Science Centre ............... 45 2.3 Svalbard in general ........................ 12 6.2.5 Land development in Longyearbyen ................................ 46 3 Framework under international 6.2.6 Energy supply ................................ 46 law .................................................... 17 6.2.7 Water supply .................................. 47 3.1 Norwegian sovereignty .................. 17 6.3 Provision of services ..................... -

Prioriterte Kulturminner Og Kulturmiljøer Pa Svalbard
KATALOG PRIORITERTE KULTURMINNER OG KULTURMILJØER PA SVALBARD Versjon 1.1 (2013) Irene Skauen Sandodden Sysselmannen på Svalbard Katalog prioriterte kulturminner og kulturmiljøer på Svalbard, versjon 1.1 Side 1 Telefon 79 02 43 00 Internett Adresse Telefaks 79 02 11 66 www.sysselmannen.no Sysselmannen på Svalbard, E-post [email protected] Pb. 633, 9171 Longyearbyen ISBN: Tilgjengelighet Internett: www.sysselmannen.no Opplag: Trykkes ikke, kun digitalt Utgiver Årstall: 2013 Sysselmannen på Svalbard, miljøvernavdelingen Sider: 220 Forfattere Irene Skauen Sandodden. Tekt er hentet fra ulike kilder. Per Kyrre Reymert, Tora Hultgreen, Marit Anne Hauan og Thor Bjørn Arlov har skrevet artikler om de ulike fasene i Svalbard historie (kapittel 2). Deltakende institusjoner Sysselmannen på Svalbard Tittel Title Katalog prioriterte kulturminner og kulturmiljøer på Svalbard. Versjon Catalogue of the cultural heritage sites with high priority in Svalbard. 1.1 (2013). Version 1.1 (2013). Referanse Katalog prioriterte kulturminner og kulturmiljøer på Svalbard. Versjon 1.1 (2013). Tilgjengelig på Internett: www.sysselmannen.no. Sammendrag Katalogen gir et kort innblikk i historien til Svalbard og representative kulturminner. Videre beskrives de om lag 100 prioriterte kulturminnene og kulturmiljøene som er valgt ut i Kulturminneplan for Svalbard 2013 – 2023. Katalogen er utarbeidet som et vedlegg til kulturminneplanen, men kan revideres ved behov. Emneord norsk Keywords English - Kulturminner og kulturmiljø - Cultural heritage (monuments and cultural -

Late Quaternary Environmental History of Central Prins Karls Forland, Western Svalbard
Late Quaternary environmental history of central Prins Karls Forland, western Svalbard TORBJO¨ RN ANDERSSON, STEVEN L. FORMAN, O´ LAFUR INGO´ LFSSON AND WILLIAM F. MANLEY Andersson, T., Forman, S. L., Ingo´lfsson, O´ . & Manley, W. F. 1999 (June): Late Quaternary environmental history of central Prins Karls Forland, western Svalbard. Boreas, Vol. 28, pp. 292–307. Oslo. ISSN 0300-9483. This paper presents the results from stratigraphic and geomorphologic investigations in the Poolepynten area, Prins Karls Forland, western Svalbard. Field mapping, soil profile development and 14C dating reveal the exis- tence of at least two generations of raised beach deposits. Well-developed raised beaches rise to the Late Weichselian marine limit at 36 m a.s.l. Discontinuous pre-Late Weichselian beach deposits rise from the Late Weichselian marine limit to approximately 65 m a.s.l. Expansion of local glaciers in the area during the Late Weichselian is indicated by a till that locally overlies pre-Late Weichselian raised beach deposits. Strati- graphic data from coastal sections reveal two shallow marine units deposited during part of oxygen isotope stage 5. The two shallow marine units are separated by a subglacially deposited till that indicates an ice ad- vance from Prins Karls Forland into the Forlandsundet basin some time during the latter part of stage 5. Dis- continuous glaciofluvial deposits and a cobble-boulder lag could relate to a Late Weichselian local glacial ad- vance across the coastal site. Late Weichselian/early Holocene beach deposits cap the sedimentary succession. Palaeotemperature estimates derived from amino acid ratios in subfossil marine molluscs indicate that the area has not been submerged or covered by warm based glacier ice for significant periods of time during the time interval ca.70kato10ka. -

Landvik Etal 2014 QSR.Pdf
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/authorsrights Author's personal copy Quaternary Science Reviews 92 (2014) 258e268 Contents lists available at ScienceDirect Quaternary Science Reviews journal homepage: www.elsevier.com/locate/quascirev Landscape imprints of changing glacial regimes during ice-sheet build-up and decay: a conceptual model from Svalbard Jon Y. Landvik a,*, Helena Alexanderson b, Mona Henriksen a, Ólafur Ingólfsson c,d a Department of Environmental Sciences, Norwegian University of Life Sciences, P.O. Box 5003, NO-1432 Ås, Norway b Department of Geology, Lund University, Sölvegatan 12, SE-223 62 Lund, Sweden c Faculty of Earth Sciences, University of Iceland, Sturlugata 7, IS-101 Reykjavik, Iceland d University Centre in Svalbard, UNIS, P.O. Box 156, NO-9171 Longyearbyen, Norway article info abstract Article history: The behaviour of ice sheets and their geologic imprints in fjord regions are often multifaceted. Fjords, Received 13 February 2013 which were temporarily occupied by fast flowing ice-streams during major glaciations, and inter-fjord Received in revised form areas, which were covered by less active ice, show different signatures of past glaciations. -

Reindeer on Prins Karls Forland, Svalbard
Research Note Reindeer on Prins Karls Forland, Svalbard IAN GJERTZ Gjertz, I. 1995: Reindeer on Prins Karls Forland, Svalbard 1995. Polar Research 14(1). 87-88. Svalbard reindeer were exterminated on the island of Prins Karls Forland just after the turn of the last century. In July 1994, after a period of 90 years, reindeer were again observed on the island. Ian Gjertz, Norsk Polarinstitutt, P. 0. Box 5072 Majorstuen, N-0301 Oslo, Norway. The Svalbard subspecies of reindeer (Rangifer On Prins Karls Forland reindeer were appar- tarandus platyrhynchus) is only found in the Sval- ently exterminated before 1926 (Wollebzk 1926). bard archipelago. When the archipelago was dis- According to Bruce (1908), who mapped Prins covered, around year 1600, reindeer were Karls Forland just after the turn of the century, abundant throughout much of the ice-free parts reindeer were occasionally found on the island. of Svalbard (Lon0 1959). Since then, excessive In 1906 Bruce (1907) saw two reindeer at the hunting reduced the numbers and distribution northern end of the island. Further south on the dramatically. According to Wollebaek (1926) island there were two groups of overwintering hunting drove the reindeer from the easily access- trappers in 1907. The leader of one of these ible western coast of Spitsbergen, while they were groups kept a detailed diary in which all game still abundant on the northern and eastern coasts caught or seen were registered (Pedersen 1908); of Spitsbergen as well as on Barents~yaand Edg- however, no mention is made of reindeer. This e~ya.The original size of the reindeer population suggests that reindeer must have been scarce. -
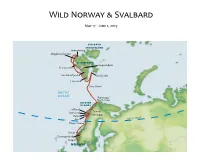
2019 Wild Norway & Svalbard Field Report
Wild Norway & Svalbard May 17 - June 2, 2019 SVALBARD ARCHIPELAGO Smeerenburg Magdalenefjorden SPITSBERGEN Longyearbyen Poolepynten Van Mijenfjorden Storfjorden Hornsund Bear Island ARCTIC OCEAN Skarsvaag/ North Cape LOFOTEN ISLANDS Trollfjord Stamsund Tromsø Kjerringøy Reine CLE Røst ARCTIC CIR Husey/Sanna Runde Geirangerfjord Bergen NORWAY Sunday, May 19, 2019 Bergen, Norway / Embark Ocean Adventurer Unusual for Bergen, it was a dry, warm, and sunny weekend when we arrived here to begin our travels in Wild Norway and Svalbard. Bergen experiences only five rain-free days a year, and this exceptional weather was being fully enjoyed by the locals, especially as this was also the Norwegian National Day holiday weekend. We set out on foot early this morning, for a tour of the old wharf-side district of Bryggen with its picturesque and higgledy-piggledy medieval wooden houses, their colorful gables lit up by today’s bright sunshine. Next, we boarded coaches which took us to Troldhaugen, the home of Norwegian composer Edvard Grieg for over 20 years. The hut where he did much of his work still stands on the lakeside at the foot of the garden. We were treated to an excellent piano recital of some of Grieg’s music before we went to lunch. All too soon, it was time to return to Bergen where our home for the next two weeks, the Ocean Adventurer, awaited us. Once settled in our cabins, our Expedition Leader, John Yersin, introduced us to his team of Zodiac drivers and lecturers who would be accompanying us on our journey over the next two weeks. -

Polar-Arctic-11-23Ju
Your next travel bucketlist ! POLAR ARCTIC SVALBARD 2021 Polar Experience “Enjoy the immense beauty of Svalbard on this Arctic adventure cruise among whales, walruses, polar bears and millions of sea birds. Experience high summer in the Arctic with Ocean Atlantic - one of the few ice-class expedition ships built to withstand the North Pole’s pack ice. Experience some of the Arctic’s best adventure, learn about a rich history and culture and witness 24-hour daylight under the midnight sun.” Cruise -Map Route PAGE 2 POLAR ARCTIC SVALBARD 2021 Itinerary Detail Day 1- 11 July Depart from USA You depart day from USA will be on 11 July to arrive in Oslo on 12July Overnight in the plane Day 2- 12 July Arrival Oslo Arrive at Oslo airport Overnight at Hotel in Oslo Day 3 - 13 July Departure from Oslo to Svalbard: LONGYEARBYEN, SPITSBERGEN. EMBARK THE OCEAN ATLANTIC Arrival to Longyearbyen, Capital of Svalbard – possibly the northernmost ‘real’ town in the world. Our vessel, Ocean Atlantic, is docked close to the town center. After boarding and a welcome drink, the Expedition Leader will provide information about the voyage, the ship's daily routines and the various security and safety procedures. Before sailing, there will be a mandatory safety drill. The Captain then takes the ship out of Advent Fjord and our Arctic adventure commences. Day 4- 14 July NY ÅLESUND, NY LONDON AND LILIENHÖÖK GLACIER During the ‘night’ (what is night, when the sun never sets?), we have passed Prins Karls Forland and have arrived in the magnificent Kongsfjord.