Demographic Variation of Wolbachia Infection in the Endangered Mitchell’S Satyr Butterfly
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Natural History and Conservation Genetics of the Federally Endangered Mitchell’S Satyr Butterfly, Neonympha Mitchellii Mitchellii
NATURAL HISTORY AND CONSERVATION GENETICS OF THE FEDERALLY ENDANGERED MITCHELL’S SATYR BUTTERFLY, NEONYMPHA MITCHELLII MITCHELLII By Christopher Alan Hamm A DISSRETATION Submitted to Michigan State University in partial fulfillment of the requirements for the degree of DOCTOR OF PHILOSOPHY Entomology Ecology, Evolutionary Biology and Behavior – Dual Major 2012 ABSTRACT NATURAL HISTORY AND CONSERVATION GENETICS OF THE FEDERALLY ENDANGERED MITCHELL’S SATYR BUTTERFLY, NEONYMPHA MITCHELLII MITCHELLII By Christopher Alan Hamm The Mitchell’s satyr butterfly, Neonympha mitchellii mitchellii, is a federally endangered species with protected populations found in Michigan, Indiana, and wherever else populations may be discovered. The conservation status of the Mitchell’s satyr began to be called into question when populations of a phenotypically similar butterfly were discovered in the eastern United States. It is unclear if these recently discovered populations are N. m. mitchellii and thus warrant protection. In order to clarify the conservation status of the Mitchell’s satyr I first acquired sample sizes large enough for population genetic analysis I developed a method of non- lethal sampling that has no detectable effect on the survival of the butterfly. I then traveled to all regions in which N. mitchellii is known to be extant and collected genetic samples. Using a variety of population genetic techniques I demonstrated that the federally protected populations in Michigan and Indiana are genetically distinct from the recently discovered populations in the southern US. I also detected the presence of the reproductive endosymbiotic bacterium Wolbachia, and surveyed addition Lepidoptera of conservation concern. This survey revealed that Wolbachia is a real concern for conservation managers and should be addressed in management plans. -

Designation of a Neotype for Mitchellâ•Žs Satyr, Neonympha Mitchellii
The Great Lakes Entomologist Volume 40 Numbers 3 & 4 - Fall/Winter 2007 Numbers 3 & Article 11 4 - Fall/Winter 2007 October 2007 Designation of a Neotype for Mitchell’s Satyr, Neonympha Mitchellii (Lepidoptera: Nymphalidae) Christopher A. Hamm Michigan State University Follow this and additional works at: https://scholar.valpo.edu/tgle Part of the Entomology Commons Recommended Citation Hamm, Christopher A. 2007. "Designation of a Neotype for Mitchell’s Satyr, Neonympha Mitchellii (Lepidoptera: Nymphalidae)," The Great Lakes Entomologist, vol 40 (2) Available at: https://scholar.valpo.edu/tgle/vol40/iss2/11 This Peer-Review Article is brought to you for free and open access by the Department of Biology at ValpoScholar. It has been accepted for inclusion in The Great Lakes Entomologist by an authorized administrator of ValpoScholar. For more information, please contact a ValpoScholar staff member at [email protected]. Hamm: Designation of a Neotype for Mitchell’s Satyr, <i>Neonympha Mitch 2007 THE GREAT LAKES ENTOMOLOGIST 201 DESIGNATION OF A NEOTYPE FOR MITCHELL’S SATYR, Neonympha miTchellii (Lepidoptera: Nymphalidae) Christopher A. Hamm1 The Mitchell’s satyr, Neonympha mitchellii French 1889 (Lepidoptera: Nymphalidae) was described as a new species based on a series of six males and four females collected by J. N. Mitchell from “Wakelee bog” in Cass County, Michigan (French 1889). French did not designate a holotype from this series. Much of French’s collection, and the original material included in the description, are thought to be lost (J. Shuey, M. Nielsen and J. Wilker, pers. comm.). I did not find the syntype series ofNeonympha mitchellii in potential re- positories including the collections of the American Museum of Natural History, Michigan State University, the University of Michigan, and the Field Museum of Natural History. -

Insect Survey of Four Longleaf Pine Preserves
A SURVEY OF THE MOTHS, BUTTERFLIES, AND GRASSHOPPERS OF FOUR NATURE CONSERVANCY PRESERVES IN SOUTHEASTERN NORTH CAROLINA Stephen P. Hall and Dale F. Schweitzer November 15, 1993 ABSTRACT Moths, butterflies, and grasshoppers were surveyed within four longleaf pine preserves owned by the North Carolina Nature Conservancy during the growing season of 1991 and 1992. Over 7,000 specimens (either collected or seen in the field) were identified, representing 512 different species and 28 families. Forty-one of these we consider to be distinctive of the two fire- maintained communities principally under investigation, the longleaf pine savannas and flatwoods. An additional 14 species we consider distinctive of the pocosins that occur in close association with the savannas and flatwoods. Twenty nine species appear to be rare enough to be included on the list of elements monitored by the North Carolina Natural Heritage Program (eight others in this category have been reported from one of these sites, the Green Swamp, but were not observed in this study). Two of the moths collected, Spartiniphaga carterae and Agrotis buchholzi, are currently candidates for federal listing as Threatened or Endangered species. Another species, Hemipachnobia s. subporphyrea, appears to be endemic to North Carolina and should also be considered for federal candidate status. With few exceptions, even the species that seem to be most closely associated with savannas and flatwoods show few direct defenses against fire, the primary force responsible for maintaining these communities. Instead, the majority of these insects probably survive within this region due to their ability to rapidly re-colonize recently burned areas from small, well-dispersed refugia. -

Integrated Management Guidelines for Four Habitats and Associated
Integrated Management Guidelines for Four Habitats and Associated State Endangered Plants and Wildlife Species of Greatest Conservation Need in the Skylands and Pinelands Landscape Conservation Zones of the New Jersey State Wildlife Action Plan Prepared by Elizabeth A. Johnson Center for Biodiversity and Conservation American Museum of Natural History Central Park West at 79th Street New York, NY 10024 and Kathleen Strakosch Walz New Jersey Natural Heritage Program New Jersey Department of Environmental Protection State Forestry Services Office of Natural Lands Management 501 East State Street, 4th Floor MC501-04, PO Box 420 Trenton, NJ 08625-0420 For NatureServe 4600 N. Fairfax Drive – 7th Floor Arlington, VA 22203 Project #DDF-0F-001a Doris Duke Charitable Foundation (Plants) June 2013 NatureServe # DDCF-0F-001a Integrated Management Plans for Four Habitats in NJ SWAP Page | 1 Acknowledgments: Many thanks to the following for sharing their expertise to review and discuss portions of this report: Allen Barlow, John Bunnell, Bob Cartica, Dave Jenkins, Sharon Petzinger, Dale Schweitzer, David Snyder, Mick Valent, Sharon Wander, Wade Wander, Andy Windisch, and Brian Zarate. This report should be cited as follows: Johnson, Elizabeth A. and Kathleen Strakosch Walz. 2013. Integrated Management Guidelines for Four Habitats and Associated State Endangered Plants and Wildlife Species of Greatest Conservation Need in the Skylands and Pinelands Landscape Conservation Zones of the New Jersey State Wildlife Action Plan. American Museum of Natural History, Center for Biodiversity and Conservation and New Jersey Department of Environmental Protection, Natural Heritage Program, for NatureServe, Arlington, VA. 149p. NatureServe # DDCF-0F-001a Integrated Management Plans for Four Habitats in NJ SWAP Page | 2 TABLE OF CONTENTS Page Project Summary………………………………………………………………………………………..…. -

Executive Summary
A Guide to the Natural Communities of the Delaware Estuary June 2006 Citation: Westervelt, K., E. Largay, R. Coxe, W. McAvoy, S. Perles, G. Podniesinski, L. Sneddon, and K. Strakosch Walz. 2006. A Guide to the Natural Communities of the Delaware Estuary: Version 1. NatureServe. Arlington, Virginia. PDE Report No. 06-02 Copyright © 2006 NatureServe COVER PHOTOS Top L: Eastern Hemlock - Great Laurel Swamp, photo from Pennsylvania Natural Heritage Top R: Pitch Pine - Oak Forest, photo by Andrew Windisch, photo from New Jersey Natural Heritage Bottom R: Maritime Red Cedar Woodland, photo by Robert Coxe, photo from Delaware Natural Heritage Bottom L: Water Willow Rocky Bar and Shore in Pennsylvania, photo from Pennsylvania Natural Heritage A GUIDE TO THE NATURAL COMMUNITIES OF THE DELAWARE ESTUARY Kellie Westervelt Ery Largay Robert Coxe William McAvoy Stephanie Perles Greg Podniesinski Lesley Sneddon Kathleen Strakosch Walz. Version 1 June 2006 TABLE OF CONTENTS PREFACE ................................................................................................................................11 ACKNOWLEDGEMENTS ............................................................................................................. 12 INTRODUCTION........................................................................................................................ 13 CLASSIFICATION APPROACH..................................................................................................... 14 International Terrestrial Ecological Systems Classification -

Tympanal Ears in Nymphalidae Butterflies: Morphological Diversity and Tests on the Function of Hearing
Tympanal Ears in Nymphalidae Butterflies: Morphological Diversity and Tests on the Function of Hearing by Laura E. Hall A thesis submitted to the Faculty of Graduate Studies and Postdoctoral Affairs in partial fulfillment of the requirements for the degree of Master of Science in Biology Carleton University Ottawa, Ontario, Canada © 2014 Laura E. Hall i Abstract Several Nymphalidae butterflies possess a sensory structure called the Vogel’s organ (VO) that is proposed to function in hearing. However, little is known about the VO’s structure, taxonomic distribution or function. My first research objective was to examine VO morphology and its accessory structures across taxa. Criteria were established to categorize development levels of butterfly VOs and tholi. I observed that enlarged forewing veins are associated with the VOs of several species within two subfamilies of Nymphalidae. Further, I discovered a putative light/temperature-sensitive organ associated with the VOs of several Biblidinae species. The second objective was to test the hypothesis that insect ears function to detect bird flight sounds for predator avoidance. Neurophysiological recordings collected from moth ears show a clear response to flight sounds and chirps from a live bird in the laboratory. Finally, a portable electrophysiology rig was developed to further test this hypothesis in future field studies. ii Acknowledgements First and foremost I would like to thank David Hall who spent endless hours listening to my musings and ramblings regarding butterfly ears, sharing in the joy of my discoveries, and comforting me in times of frustration. Without him, this thesis would not have been possible. I thank Dr. -

Mitchell's Satyr Butterfly, Neonympha Mitchellii Mitchellii French, in Southwestern Michigan
Mitchell’s Satyr Programmatic Safe Harbor Agreement This Programmatic Safe Harbor Agreement, effective and binding on the date of last signature below, is between U.S. Fish and Wildlife Service’s East Lansing Field Office Project Leader and the U.S. Fish and Wildlife Service. Permittee: Scott Hicks, Project Leader U.S. Fish and Wildlife Service East Lansing Field Office 2651 Coolidge Road, Suite 101 East Lansing, Michigan 48823 (517) 351-2555 The Service designates the following as the Agreement Contact: Laura Ragan, Recovery Coordinator U.S. Fish & Wildlife Service, Region 3 5600 American Blvd. West, Suite 990 Bloomington, Minnesota 55437-1458 Tracking Number: Summary of Purpose of the SHA: The purpose of this agreement is to outline conservation actions that participating property owners will implement and monitor on their enrolled properties for Mitchell’s satyr (Neonympha mitchellii mitchellii). The goal of the agreement is to encourage property owners to engage in conservation actions for the Mitchell’s satyr that provide a net conservation benefit to recovery. 1.0 Introduction The U.S. Fish and Wildlife Service (Service) Safe Harbor Program (64 FR 32717) provides regulatory flexibility to non-federal landowners who voluntarily commit to implementing or avoiding specific activities, over a defined timeframe, that are reasonably expected to provide a net conservation benefit to species listed under the Endangered Species Act of 1973, as amended (Act). In exchange for this commitment, enrolled landowners (Cooperators) receive assurances from the Service that no additional future regulatory restrictions will be imposed or commitments required for species covered under a Safe Harbor Agreement. -

Habitat Characterization of Five Rare Insects in Michigan (Lepidoptera: Hesperiidae, Riodinidae, Satyridae; Homoptera: Cercopidae)
The Great Lakes Entomologist Volume 32 Number 3 - Fall 1999 Number 3 - Fall 1999 Article 13 October 1999 Habitat Characterization of Five Rare Insects in Michigan (Lepidoptera: Hesperiidae, Riodinidae, Satyridae; Homoptera: Cercopidae) Keith S. Summerville Miami University Christopher A. Clampitt The Nature Conservancy Follow this and additional works at: https://scholar.valpo.edu/tgle Part of the Entomology Commons Recommended Citation Summerville, Keith S. and Clampitt, Christopher A. 1999. "Habitat Characterization of Five Rare Insects in Michigan (Lepidoptera: Hesperiidae, Riodinidae, Satyridae; Homoptera: Cercopidae)," The Great Lakes Entomologist, vol 32 (2) Available at: https://scholar.valpo.edu/tgle/vol32/iss2/13 This Peer-Review Article is brought to you for free and open access by the Department of Biology at ValpoScholar. It has been accepted for inclusion in The Great Lakes Entomologist by an authorized administrator of ValpoScholar. For more information, please contact a ValpoScholar staff member at [email protected]. Summerville and Clampitt: Habitat Characterization of Five Rare Insects in Michigan (Lepido 1999 THE GREAT LAKES ENTOMOLOGIST 225 HABITAT CHARACTERIZATION OF FIVE RARE INSECTS IN MICHIGAN (LEPIDOPTERA: HESPERIIDAE, RIODINIDAE, SATYRIDAE; HOMOPTERA: CERCOPIDAE) Keith S. Summerville 1,2 and Christopher A. Clampitt! ABSTRACT Over 80 species ofinsects are listed as endangered, threatened, or special concern under Michigan's endangered species act. For the majority of these species, detailed habitat information is scant or difficult to interpret. We de scribe the habitat of five insect species that are considered rare in Michigan: Lepyronia angulifera (Cercopidae), Prosapia ignipectus (Cercopidae), Oarisma poweshiek (Hesperiidae), Calephelis mutica (Riodinidae), and Neonympha mitchellii mitchellii (Satyridae). Populations of each species were only found within a fraction of the plant communities deemed suitable based upon previous literature. -

ABSTRACT a Thorough Understanding of the Mechanisms Driving Larger Scale Consequences of Movement First Requires an Understandin
ABSTRACT Kuefler, Daniel Cory. Local versus landscape determinants of butterfly movement behaviors. (Under the direction of Nick M. Haddad) A thorough understanding of the mechanisms driving larger scale consequences of movement first requires an understanding of whether movement behaviors are related to local or landscape scale determinants. I studied the movement behaviors of four species of bottomland-dwelling butterflies in a natural setting to examine the determinants of movement behavior across different scales. Across spatial scales, I tested the relative importance and predictive value of three landscape attributes: topography, boundary contrast, and stream proximity, and two local habitat attributes: host plant cover and comprehensive vegetative structure. Across species, I tested the relative importance of organism size and habitat specificity to explain response variation. In general, butterfly responses to landscape features were stronger and more universal while responses to local features were weaker and more variable by species. Specifically, results from this study showed that topography does not influence movement behaviors but boundary contrast, stream proximity, and host plant abundance all contributed to movement patterns. Orientation to these features was not related to organism size, but did vary in accordance with habitat specificity. These results suggest that studies on dispersal in fragmented landscapes should consider the effects of that fragmentation on multiple scales. This consideration is particularly important in the management of rare species, when specific behaviors may ultimately affect the success of conservation efforts. LOCAL VERSUS LANDSCAPE DETERMINANTS OF BUTTERFLY MOVEMENT BEHAVIORS By DANIEL CORY KUEFLER A thesis submitted to the graduate faculty of North Carolina State University in partial fulfillment of the requirements for the degree of Master of Science ZOOLOGY Raleigh 2005 Approved by: ______________________________, ______________________________ Christopher E. -
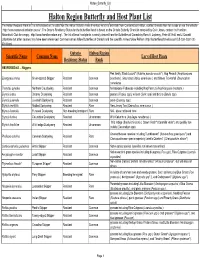
Halton Region Butterfly and Host Plant List
Halton_Butterfly_List Halton Region Butterfly and Host Plant List The Halton Regional Rank in this list is based on the data from the Halton Natural Areas Inventory which is available from Conservation Halton, contact Brenda Axon for a copy or see the website : http://www.conservationhalton.on.ca/ . The Ontario Residency Status for the butterflies listed is based on the Ontario Butterfly Checklist compiled by Colin Jones, contact the Hamilton Naturalists' Club for a copy : http://www.hamiltonnature.org/ . The list of larval host plants is mostly derived from the Butterflies of Canada by Ross A. Layberry, Peter W. Hall, and J. Donald Lafontaine but other sources may have been referenced. Common names follow Butterflies of Canada and the scientific names follow Pelham (http://butterfliesofamerica.com/US-Can-Cat-1-30- 2011.htm). Ontario Halton Region Scientific Name Common Name Larval Host Plants Residency Status Rank HESPERIIDAE – Skippers Pea family; Black Locust* (Robinia pseudo-acacia* ), Hog Peanut (Amphicarpaea Epargyreus clarus Silver-spotted Skipper Resident Common bracteata ), Groundnut (Apios americana ), and Showy Tick-trefoil (Desmodium canadense ) Thorybes pylades Northern Cloudywing Resident Common herbaciouse Fabaceae including Hog Peanut (Amphicarpaea bracteata ) Erynnis icelus Dreamy Duskywing Resident Common poplars (Poplus spp), willows (Salix spp) and birchs (Betula spp) Erynnis juvenalis Juvenal's Duskywing Resident Common oaks (Quercus spp) Erynnis martialis Mottled Duskywing Resident Rare New Jersey Tea (Ceanothus -

Comprehensive Survey of Butterflies at the Sam D. Hamilton Noxubee National Wildlife Refuge
Mississippi State University Scholars Junction Theses and Dissertations Theses and Dissertations 12-1-2020 Comprehensive Survey of Butterflies at the Sam .D Hamilton Noxubee National Wildlife Refuge Jordan Gesell Follow this and additional works at: https://scholarsjunction.msstate.edu/td Recommended Citation Gesell, Jordan, "Comprehensive Survey of Butterflies at the Sam .D Hamilton Noxubee National Wildlife Refuge" (2020). Theses and Dissertations. 1130. https://scholarsjunction.msstate.edu/td/1130 This Graduate Thesis - Open Access is brought to you for free and open access by the Theses and Dissertations at Scholars Junction. It has been accepted for inclusion in Theses and Dissertations by an authorized administrator of Scholars Junction. For more information, please contact [email protected]. Template B v4.1 (beta): Created by L. Threet 11/15/19 Comprehensive survey of butterflies at the Sam D. Hamilton Noxubee National Wildlife Refuge By TITLE PAGE Jordan Lambert Gesell Approved by: JoVonn G. Hill (Major Professor) Brandon Barton Jeffrey W. Harris Kenneth Willeford (Graduate Coordinator) Scott T. Willard (Dean, College of Agriculture and Life Sciences) A Thesis Submitted to the Faculty of Mississippi State University in Partial Fulfillment of the Requirements for the Degree of Master of Science in Agricultural Life Sciences in the Department of Biochemistry, Molecular Biology, Entomology and Plant Pathology Mississippi State, Mississippi November 2020 Copyright by COPYRIGHT PAGE Jordan Lambert Gesell 2020 Name: Jordan Lambert Gesell ABSTRACT Date of Degree: November 25, 2020 Institution: Mississippi State University Major Field: Agricultural Life Sciences Major Professor: JoVonn G. Hill Title of Study: Comprehensive survey of butterflies at the Sam D. -

Sentinels on the Wing: the Status and Conservation of Butterflies in Canada
Sentinels on the Wing The Status and Conservation of Butterflies in Canada Peter W. Hall Foreword In Canada, our ties to the land are strong and deep. Whether we have viewed the coasts of British Columbia or Cape Breton, experienced the beauty of the Arctic tundra, paddled on rivers through our sweeping boreal forests, heard the wind in the prairies, watched caribou swim the rivers of northern Labrador, or searched for song birds in the hardwood forests of south eastern Canada, we all call Canada our home and native land. Perhaps because Canada’s landscapes are extensive and cover a broad range of diverse natural systems, it is easy for us to assume the health of our important natural spaces and the species they contain. Our country seems so vast compared to the number of Canadians that it is difficult for us to imagine humans could have any lasting effect on nature. Yet emerging science demonstrates that our natural systems and the species they contain are increas- ingly at risk. While the story is by no means complete, key indicator species demonstrate that Canada’s natural legacy is under pressure from a number of sources, such as the conversion of lands for human uses, the release of toxic chemicals, the introduction of new, invasive species or the further spread of natural pests, and a rapidly changing climate. These changes are hitting home and, with the globalization and expansion of human activities, it is clear the pace of change is accelerating. While their flights of fancy may seem insignificant, butterflies are sentinels or early indicators of this change, and can act as important messengers to raise awareness.