The Growth and Duration of Life of Celosia Cris- Tata Seedlings at Different Temperatures
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Yearbook of Agriculture • 1961 ^
87TH CONGRESS, IST SESSION, HOUSE DOCUMENT NO. 29 THE YEARBOOK OF AGRICULTURE • 1961 ^ THE UNITED STATES DEPARTMENT OF AGRICULTURE Washington, D.G. SEEDS The Yearbook of Agriculture 1961 ¿^ f i Í THE UNITED STATES GOVERNMENT PRINTING OFFICE FOR SALE BY THE SUPERINTENDENT OF DOCUMENTS, WASHINGTON 25, D.G., PRICE $2 FOREWORD ORVILLE L. FREEMAN Secretary of Agriculture GOOD SEEDS ARE both a symbol and a foundation of the good Ufe our people have gained. A baisic factor in our realization of mankind's most sought goal, agricultural abundance, good seeds can be a means of our bringing about an Age of Plenty and an Age of Peace and Free- dom. We can use our good seeds to help end hunger and fear for the less fortunate half of the human family. So used, our seeds can be more meaningful to a hungry world than can the rocket that first carries man to the moon. This Yearbook of Agriculture seeks to provide a new and improved basis for understanding the complex order of Nature's forces so that man can better shape them in a positive and creative fashion. Seeds are ever a positive and creative force. Seeds are the germ of life, a beginning and an end, the fruit of yesterday's harvest and the promise of tomorrow's. Without an ample store of seeds there can be no national treasure, or no future for a Nation. Finding and developing better seeds is the oldest continuous service our Federal Government has rendered to our farmers—indeed, to all our people. -
S. Frugiperda (J
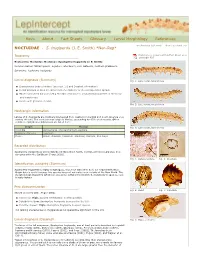
Keys About Fact Sheets Glossary Larval Morphology References << Previous fact sheet Next fact sheet >> NOCTUIDAE - S. frugiperda (J. E. Smith) *Non-Rep* Taxonomy Click here to download this Fact Sheet as a printable PDF Noctuoidea: Noctuidae: Noctuinae: Spodoptera frugiperda (J. E. Smith) Common names: fall armyworm, cogollero, whorlworm, corn leafworm, southern grassworm Synonyms: Laphygma frugiperda Larval diagnosis (Summary) Fig. 1: Late instar, lateral view Characteristic body coloration (see Figs. 1-3 and Detailed Information) Dorsal pinacula of A1-8 are larger than the diameter of the corresponding spiracle Minute sclerotized bar connecting the SD1 setal base to a tonofibrillary platelet on the meso- and metathorax Cuticle with granulose texture Fig. 2: Late instar, lateral view Host/origin information Larvae of S. frugiperda are routinely intercepted from locations in Central and South America on a variety of hosts. The most common origin is Mexico, accounting for 65% of all records. Other common origin/host combinations are listed here: Origin Host(s) Fig. 3: Late instar, lateral view Colombia Alstroemeria, Chrysanthemum, Gerbera Dominican Republic Capsicum Mexico Apium, Brassica, Capsicum, Gladiolus, Ocimum, Zea mays Recorded distribution Spodoptera frugiperda is widely distributed throughout North, Central, and South America. It is also present in the Caribbean (Pogue 2002). Fig. 4: Cuticle texture Fig. 5: Crochets Identifcation authority (Summary) Spodoptera frugiperda is highly polyphagous, thus host data often does not help identification. Origin data is useful because this species does not naturally occur outside of the New World. The morphological characters listed here should be sufficient to identify S. frugiperda to species, even in early instars. -

Dictionary of Cultivated Plants and Their Regions of Diversity Second Edition Revised Of: A.C
Dictionary of cultivated plants and their regions of diversity Second edition revised of: A.C. Zeven and P.M. Zhukovsky, 1975, Dictionary of cultivated plants and their centres of diversity 'N -'\:K 1~ Li Dictionary of cultivated plants and their regions of diversity Excluding most ornamentals, forest trees and lower plants A.C. Zeven andJ.M.J, de Wet K pudoc Centre for Agricultural Publishing and Documentation Wageningen - 1982 ~T—^/-/- /+<>?- •/ CIP-GEGEVENS Zeven, A.C. Dictionary ofcultivate d plants andthei rregion so f diversity: excluding mostornamentals ,fores t treesan d lowerplant s/ A.C .Zeve n andJ.M.J ,d eWet .- Wageninge n : Pudoc. -11 1 Herz,uitg . van:Dictionar y of cultivatedplant s andthei r centreso fdiversit y /A.C .Zeve n andP.M . Zhukovsky, 1975.- Me t index,lit .opg . ISBN 90-220-0785-5 SISO63 2UD C63 3 Trefw.:plantenteelt . ISBN 90-220-0785-5 ©Centre forAgricultura l Publishing and Documentation, Wageningen,1982 . Nopar t of thisboo k mayb e reproduced andpublishe d in any form,b y print, photoprint,microfil m or any othermean swithou t written permission from thepublisher . Contents Preface 7 History of thewor k 8 Origins of agriculture anddomesticatio n ofplant s Cradles of agriculture and regions of diversity 21 1 Chinese-Japanese Region 32 2 Indochinese-IndonesianRegio n 48 3 Australian Region 65 4 Hindustani Region 70 5 Central AsianRegio n 81 6 NearEaster n Region 87 7 Mediterranean Region 103 8 African Region 121 9 European-Siberian Region 148 10 South American Region 164 11 CentralAmerica n andMexica n Region 185 12 NorthAmerica n Region 199 Specieswithou t an identified region 207 References 209 Indexo fbotanica l names 228 Preface The aimo f thiswor k ist ogiv e thereade r quick reference toth e regionso f diversity ofcultivate d plants.Fo r important crops,region so fdiversit y of related wild species areals opresented .Wil d species areofte nusefu l sources of genes to improve thevalu eo fcrops . -

Revisiting the Status of Cultivated Plant Species Agrobiodiversity in India: an Overview ANURUDH K SINGH* 2924, Sector-23, Gurgaon, Haryana, India 122 017
Proc Indian Natn Sci Acad 83 No. 1 March 2017 pp. 151-174 Printed in India. DOI: 10.16943/ptinsa/2016/v82/48406 Review Article Revisiting the Status of Cultivated Plant Species Agrobiodiversity in India: An Overview ANURUDH K SINGH* 2924, Sector-23, Gurgaon, Haryana, India 122 017 (Received on 14 March 2016; Revised on 20 May 2016; Accepted on 16 June 2016) A revisit to the literature on cultivated plant species agrobiodiversity in India revealed that the floristic diversity is represented by 17,926 species of angiosperm, while Indian agriculture cultivates 811 plant species and harbours more than 900 wild relatives of the cultivated plant species distributed over 10 (+ 1) biogeographic regions of the the country, significantly higher than commonly cited in the literature. Further, it revealed the role of Indian communities in domestication to cultivation of around 215 economically important plant species, and adaption of around 600 exotic crop species. Based on new evidence, several species require inclusion and others deletion, and many need further investigations to resolve the issue on country of their origin. Cultivation of crop species in diverse natural and man-made agroecological systems for centuries has generated a huge amount of genetic diversity in a large number of crop species, maintained by the farmers in the form of landraces or farmer’s varieties, and conserved as collections/accessions in the national agricultural research system. Keywords: Agrobiodiversity; Domestication; Cultivation; Genetic Diversity; Wild Relatives -

Field Identification of the 50 Most Common Plant Families in Temperate Regions
Field identification of the 50 most common plant families in temperate regions (including agricultural, horticultural, and wild species) by Lena Struwe [email protected] © 2016, All rights reserved. Note: Listed characteristics are the most common characteristics; there might be exceptions in rare or tropical species. This compendium is available for free download without cost for non- commercial uses at http://www.rci.rutgers.edu/~struwe/. The author welcomes updates and corrections. 1 Overall phylogeny – living land plants Bryophytes Mosses, liverworts, hornworts Lycophytes Clubmosses, etc. Ferns and Fern Allies Ferns, horsetails, moonworts, etc. Gymnosperms Conifers, pines, cycads and cedars, etc. Magnoliids Monocots Fabids Ranunculales Rosids Malvids Caryophyllales Ericales Lamiids The treatment for flowering plants follows the APG IV (2016) Campanulids classification. Not all branches are shown. © Lena Struwe 2016, All rights reserved. 2 Included families (alphabetical list): Amaranthaceae Geraniaceae Amaryllidaceae Iridaceae Anacardiaceae Juglandaceae Apiaceae Juncaceae Apocynaceae Lamiaceae Araceae Lauraceae Araliaceae Liliaceae Asphodelaceae Magnoliaceae Asteraceae Malvaceae Betulaceae Moraceae Boraginaceae Myrtaceae Brassicaceae Oleaceae Bromeliaceae Orchidaceae Cactaceae Orobanchaceae Campanulaceae Pinaceae Caprifoliaceae Plantaginaceae Caryophyllaceae Poaceae Convolvulaceae Polygonaceae Cucurbitaceae Ranunculaceae Cupressaceae Rosaceae Cyperaceae Rubiaceae Equisetaceae Rutaceae Ericaceae Salicaceae Euphorbiaceae Scrophulariaceae -

Checklist of the Washington Baltimore Area
Annotated Checklist of the Vascular Plants of the Washington - Baltimore Area Part I Ferns, Fern Allies, Gymnosperms, and Dicotyledons by Stanwyn G. Shetler and Sylvia Stone Orli Department of Botany National Museum of Natural History 2000 Department of Botany, National Museum of Natural History Smithsonian Institution, Washington, DC 20560-0166 ii iii PREFACE The better part of a century has elapsed since A. S. Hitchcock and Paul C. Standley published their succinct manual in 1919 for the identification of the vascular flora in the Washington, DC, area. A comparable new manual has long been needed. As with their work, such a manual should be produced through a collaborative effort of the region’s botanists and other experts. The Annotated Checklist is offered as a first step, in the hope that it will spark and facilitate that effort. In preparing this checklist, Shetler has been responsible for the taxonomy and nomenclature and Orli for the database. We have chosen to distribute the first part in preliminary form, so that it can be used, criticized, and revised while it is current and the second part (Monocotyledons) is still in progress. Additions, corrections, and comments are welcome. We hope that our checklist will stimulate a new wave of fieldwork to check on the current status of the local flora relative to what is reported here. When Part II is finished, the two parts will be combined into a single publication. We also maintain a Web site for the Flora of the Washington-Baltimore Area, and the database can be searched there (http://www.nmnh.si.edu/botany/projects/dcflora). -

The Amaranth (Amaranthus Hypochondriacus) Genome: Genome, Transcriptome and Physical Map Assembly
Brigham Young University BYU ScholarsArchive Student Works 2015-06-01 The Amaranth (Amaranthus Hypochondriacus) Genome: Genome, Transcriptome and Physical Map Assembly Jared William Clouse Brigham Young University - Provo, [email protected] Follow this and additional works at: https://scholarsarchive.byu.edu/studentpub Part of the Plant Sciences Commons BYU ScholarsArchive Citation Clouse, Jared William, "The Amaranth (Amaranthus Hypochondriacus) Genome: Genome, Transcriptome and Physical Map Assembly" (2015). Student Works. 167. https://scholarsarchive.byu.edu/studentpub/167 This Peer-Reviewed Article is brought to you for free and open access by BYU ScholarsArchive. It has been accepted for inclusion in Student Works by an authorized administrator of BYU ScholarsArchive. For more information, please contact [email protected], [email protected]. The Amaranth (Amaranthus hypochondriacus) Genome: Genome, Transcriptome and Physical Map Assembly Jared William Clouse A thesis submitted to the faculty of Brigham Young University in partial fulfillment of the requirements for the degree of Master of Science P. Jeffery Maughan, Chair Eric N. Jellen Joshua A. Udall Department of Plant and Wildlife Sciences Brigham Young University June 2015 Copyright © 2015 Jared William Clouse All Rights Reserved ABSTRACT The Amaranth (Amaranthus Hypochondriacus) Genome: Genome, Transcriptome and Physical Map Assembly Jared William Clouse Department of Plant and Wildlife Sciences, BYU Master of Science Amaranthus hypochondriacus is an emerging pseudo-cereal native to the New World which has garnered increased attention in recent years due to its nutritional quality, in particular its seed protein, and more specifically its high levels of the essential amino acid lysine. It belongs to the Amaranthaceae family, is an ancient paleotetraploid that shows amphidiploid inheritance (2n=32), and has an estimated genome size of 466 Mb. -

Four Flowering Plant Species Described from Katanga (Democratic Republic of the Congo) Are Based on Specimens Collected in Guangxi, China
Blumea 66, 2021: 82–92 www.ingentaconnect.com/content/nhn/blumea RESEARCH ARTICLE https://doi.org/10.3767/blumea.2021.66.01.04 Four flowering plant species described from Katanga (Democratic Republic of the Congo) are based on specimens collected in Guangxi, China E. Robbrecht1,*, S. De Smedt1, P. Goetghebeur 2, P. Stoffelen1, F. Verloove1 Key words Abstract The original set of botanical collections of the agronomist H.A. Homblé is conserved in the herbarium BR. Homblé was one of the first collectors (1911–1913) for the flora of Katanga, Democratic Republic of the Congo. Caesalpinia bonduc Many Homblé specimens were described as taxonomic novelties; 107 tropical African plant species are named Caesalpinia homblei after him. Before his colonial career in Katanga, Homblé stayed about two years (1909–1911) in Guangxi, China. Digitaria abludens His incompletely labelled Chinese collections were erroneously considered as collected in Katanga. This supposed Digitaria polybotryoides African origin has led to confusion with regard to the identification, and even resulted in the description of four spe- Drosera insolita cies believed to be new for science. This paper presents and discusses Homblé’s collection made in Guangxi, and Drosera lunata the assumed novelties in it. Drosera insolita is a synonym of the Asian Drosera lunata, widespread from India to flora of Guangxi Australia. Three other species are new synonyms. Caesalpinia homblei is a synonym of the pantropical Caesalpinia flora of Katanga bonduc. Digitaria polybotryoides is a synonym of Digitaria abludens, a widespread species in tropical Asia. Grewia Grewia cuspidatoserrata katangensis is the only species that proved to be synonymous with an endemic species, Grewia cuspidatoserrata, Grewia katangensis only known from S Yunnan, and here reported as a new record for Guangxi. -

Weeds of Upland Rice and Upland Crops in Myanmar Sr. Scientific
Weeds of Upland Rice and Upland Crops in Myanmar Sr. Scientific Name English Name Order Family Source 1 Abutilum indicum Indian lantern Malvales Malvaceae The Distribution and flower Importance of Arthropod Pests and Weeds of Agriculture in Myanmar (H.Morris and D.F.Waterhouse) 2 Achyranthes aspera Devil’s horse Caryophyllales Amaranthaceae The Distribution and whip Importance of Arthropod Pests and Weeds of Agriculture in Myanmar (H.Morris and D.F.Waterhouse) 3 Aeschynomene - Fabales Fabaceae(Papilio The Distribution and aspera naceae) Importance of Arthropod Pests and Weeds of Agriculture in Myanmar (H.Morris and D.F.Waterhouse) 4 Aeschynomene joint vetch, Fabales Fabaceae(Papilio The Distribution and indica budda pea naceae) Importance of Arthropod Pests and Weeds of Agriculture in Myanmar (H.Morris and D.F.Waterhouse) 5 Ageratum Tropic Asterales Asteraceae The Distribution and conyzoides ageratum,goat Importance of Arthropod weed,blue top Pests and Weeds of Agriculture in Myanmar (H.Morris and D.F.Waterhouse) 6 Alternanthera alligator weed Caryophyllales Amaranthaceae The Distribution and philoxeroides Importance of Arthropod Pests and Weeds of Agriculture in Myanmar (H.Morris and D.F.Waterhouse) 7 Amaranthus spiny amaranth Caryophyllales Amaranthaceae The Distribution and spinosus Importance of Arthropod Pests and Weeds of Agriculture in Myanmar (H.Morris and D.F.Waterhouse) 8 Amaranthus viridis slender Caryophyllales Amaranthaceae The Distribution and amaranth Importance of Arthropod Pests and Weeds of Agriculture in Myanmar (H.Morris -

Reformation of Specialty Cut Flower Production for Celosia Cristata
Reformation of specialty cut flower production for Celosia cristata Cameron Zuck, University oF Minnesota (Landscape Design and Planning, Department oF Landscape Architecture) December 18, 2015 EXECUTIVE SUMMARY Celosia cristata is an interesting and increasingly popular crop in specialty cut flower production. With origins of dry, temperate climates in Africa and Asia, this herbaceous annual plant is now distributed and cultivated worldwide. C. cristata has a wide variety of cultivars available on the market including the ‘Chief Series’ and the ‘Bombay’. To become a more sustainable and energy-efficient crop in the future, one potential change in production is an overall increase on growth efficiency. By evaluating the current practices and traits of C. cristata, a new ideotype is UNIVERSITY OF MINNESOTA: Celosia cristata: Cut Flower Production 1 proposed that addresses possible genetic improvements that can create a more sustainable production of C. cristata. I. INTRODUCTION A. Study Species. Celosias are eye-catching flowers that come in many different colors and forms. The unique inflorescence of Celosia cristata have been compared to a rooster comb, a brain and elegant ribbon; whichever way they are described, they are sure to strike the attention of anyone who catches a glimpse (Cornell 2006). Celosia are versatile plants that have been used as herbal remedies, as ornamental annuals in the garden and as cut and dry flowers. Celosia cristata is still considered fairly unknown to consumers in specialty cut flower production and has potential to grow (Gilman and Howe 1999). Since C. celosia is a temperate climate species, field production is restricted in location and time. -

Everlastings (Dried Flowers)
Visit us on the Web: www.gardeninghelp.org Everlastings (Dried Flowers) While cut fresh flowers symbolize beauty and fleeting life, everlastings represent longevity and immortality. Everlastings are plants that retain their shape and color long after they have been picked and dried. Everlastings serve as a reminder of nature’s bounty and provide the home gardener with an opportunity to preserve the beauty of the growing season year-round. In Victorian times, flowers and leaves that retained their fragrance were used to perfume and adorn homes. Rose petals and sweet-scented rushes were scattered on floors, while cloth bags and filigree containers filled with aromatic leaves and flowers freshened drawers and cupboards. Today, interest in dried flowers is enjoying resurgence in popularity. Dried plant materials provide an attractive option for urban dwellers who may have little time, interest or space to grow a garden. Many commercial nurseries all over the world are growing flowers solely for drying. The Everlasting Garden Although good plants for drying are available in the wild and from florists and suppliers, you may want to grow your own. A garden of everlastings starts with careful planning. Since flowers and plant materials for drying should be picked as close as possible to their prime, your planting plan should take into account such information as the height and width of the mature plant, flower and foliage color as well as the time of bloom. The beds in your garden need to have a mixture of plants that will not be affected appreciably by the removal of flowers and foliage for drying. -

SRI LANKA AG: GCP/RAS/186/JPN Field Document No
COUNTRY REPORT ON THE STATE OF PLANT GENETIC RESOURCES FOR FOOD AND AGRICULTURE SRI LANKA AG: GCP/RAS/186/JPN Field Document No. 2007/08 FAO/GOVERNMENT COOPERATIVE PROGRAMME THE STATUS OF THE PGRFA IN SRI LANKA Department of Agriculture Compiled and Edited by Dr. D. H. Muthukuda Arachchi and Dr. P. M. Wijerathne Note by FAO This Country Report has been prepared by the national authorities in the context of the preparatory process for the Second Report on the State of World’s Plant Genetic Resources for Food and Agriculture. The Report is being made available by the Food and Agriculture Organization of the United Nations (FAO) as requested by the Commission on Genetic Resources for Food and Agriculture. However, the report is solely the responsibility of the national authorities. The information in this report has not been verified by FAO, and the opinions expressed do not necessarily represent the views or policy of FAO. The designations employed and the presentation of material in this information product do not imply the expression of any opinion whatsoever on the part of FAO concerning the legal or development status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. The mention of specific companies or products of manufacturers, whether or not these have been patented, does not imply that these have been endorsed or recommended by FAO in preference to others of a similar nature that are not mentioned. The views expressed in this information product are those of the author(s) and do not necessarily reflect the views of FAO.