Plan of Lecture Sound Filters External and Middle Ear Cochlear Structure
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Perforated Eardrum
Vinod K. Anand, MD, FACS Nose and Sinus Clinic Perforated Eardrum A perforated eardrum is a hole or rupture m the eardrum, a thin membrane which separated the ear canal and the middle ear. The medical term for eardrum is tympanic membrane. The middle ear is connected to the nose by the eustachian tube. A perforated eardrum is often accompanied by decreased hearing and occasional discharge. Paih is usually not persistent. Causes of Eardrum Perforation The causes of perforated eardrum are usually from trauma or infection. A perforated eardrum can occur: if the ear is struck squarely with an open hand with a skull fracture after a sudden explosion if an object (such as a bobby pin, Q-tip, or stick) is pushed too far into the ear canal. as a result of hot slag (from welding) or acid entering the ear canal Middle ear infections may cause pain, hearing loss and spontaneous rupture (tear) of the eardrum resulting in a perforation. In this circumstance, there may be infected or bloody drainage from the ear. In medical terms, this is called otitis media with perforation. On rare occasions a small hole may remain in the eardrum after a previously placed P.E. tube (pressure equalizing) either falls out or is removed by the physician. Most eardrum perforations heal spontaneously within weeks after rupture, although some may take up to several months. During the healing process the ear must be protected from water and trauma. Those eardrum perforations which do not heal on their own may require surgery. Effects on Hearing from Perforated Eardrum Usually, the larger the perforation, the greater the loss of hearing. -

Experimental Studies on the Function of the Stapedius Muscle Inman
EXPERIMENTAL STUDIES ON THE FUNCTION OF THE STAPEDIUS MUSCLE INMAN AKADEMISK AVHANDLING som med vederbörligt tillstånd av Medicinska fakulteten vid Umeå Universitet för vinnande av medicine doktorsgrad offentligen försvaras i Samhällsvetarhuset, sal D, lördagen den 25 maj 1974 kl. 9.15 f.m. av JOHN-ERIK ZAKRISSON med.lic. UMEÅ 1974 UMEÀ UNIVERSITY MEDICAL DISSERTATIONS No. 18 1974 From the Department of Otorhinolaryngology, University of Umeå, Umeå, Sweden and the Division of Physiological Acoustics, Department of Physiology II, Karolinska Institutet, Stockholm, Sweden EXPERIMENTAL STUDIES ON THE FUNCTION OF THE STAPEDIUS MUSCLE IN MAN BY JOHN-ERIK ZAKRISSON UMEÂ 1974 To Karin Eva and Gunilla The present thesis is based on the following papers which will be referred to in the text by the Roman numerals: I. Zakrisson, J.-E., Borg, E. & Blom, S. The acoustic impedance change as a measure of stapedius muscle activity in man. A methodological study with electromyography. Acta Otolaryng, preprint. II. Borg, E. & Zakrisson, J.-E. Stapedius reflex and monaural masking. Acta Otolaryng, preprint. III. Zakrisson, J.-E. The role of the stapedius reflex in poststimulatory audi tory fatigue. Acta Otolaryng, preprint. IV. Borg, E. & Zakrisson, J.-E. The activity of the stapedius muscle in man during vocalization. Acta Otolaryng, accepted for publication. CONTENTS ABBREVIATIONS .......................................... 8 INTRODUCTION.............................................................................................. 9 MATERIAL..................................................................................................... -

SENSORY MOTOR COORDINATION in ROBONAUT Richard Alan Peters
SENSORY MOTOR COORDINATION IN ROBONAUT 5 Richard Alan Peters 11 Vanderbilt University School of Engineering JSC Mail Code: ER4 30 October 2000 Robert 0. Ambrose Robotic Systems Technology Branch Automation, Robotics, & Simulation Division Engineering Directorate Richard Alan Peters II Robert 0. Ambrose SENSORY MOTOR COORDINATION IN ROBONAUT Final Report NASNASEE Summer Faculty Fellowship Program - 2000 Johnson Space Center Prepared By: Richard Alan Peters II, Ph.D. Academic Rank: Associate Professor University and Department: Vanderbilt University Department of Electrical Engineering and Computer Science Nashville, TN 37235 NASNJSC Directorate: Engineering Division: Automation, Robotics, & Simulation Branch: Robotic Systems Technology JSC Colleague: Robert 0. Ambrose Date Submitted: 30 October 2000 Contract Number: NAG 9-867 13-1 ABSTRACT As a participant of the year 2000 NASA Summer Faculty Fellowship Program, I worked with the engineers of the Dexterous Robotics Laboratory at NASA Johnson Space Center on the Robonaut project. The Robonaut is an articulated torso with two dexterous arms, left and right five-fingered hands, and a head with cameras mounted on an articulated neck. This advanced space robot, now dnven only teleoperatively using VR gloves, sensors and helmets, is to be upgraded to a thinking system that can find, in- teract with and assist humans autonomously, allowing the Crew to work with Robonaut as a (junior) member of their team. Thus, the work performed this summer was toward the goal of enabling Robonaut to operate autonomously as an intelligent assistant to as- tronauts. Our underlying hypothesis is that a robot can deveZop intelligence if it learns a set of basic behaviors ([.e., reflexes - actions tightly coupled to sensing) and through experi- ence learns how to sequence these to solve problems or to accomplish higher-level tasks. -

Eardrum Regeneration: Membrane Repair
OUTLINE Watch an animation at: Infographic: go.nature.com/2smjfq8 Pages S6–S7 EARDRUM REGENERATION: MEMBRANE REPAIR Can tissue engineering provide a cheap and convenient alternative to surgery for eardrum repair? DIANA GRADINARU he eardrum, or tympanic membrane, forms the interface between the outside world and the delicate bony structures Tof the middle ear — the ossicles — that conduct sound vibrations to the inner ear. At just a fraction of a millimetre thick and held under tension, the membrane is perfectly adapted to transmit even the faintest of vibrations. But the qualities that make the eardrum such a good conductor of sound come at a price: fra- gility. Burst eardrums are a major cause of conductive hearing loss — when sounds can’t pass from the outer to the inner ear. Most burst eardrums are caused by infections or trauma. The vast majority heal on their own in about ten days, but for a small proportion of people the perforation fails to heal natu- rally. These chronic ruptures cause conductive hearing loss and group (S. Kanemaru et al. Otol. Neurotol. 32, 1218–1223; 2011). increase the risk of middle ear infections, which can have serious In a commentary in the same journal, Robert Jackler, a head complications. and neck surgeon at Stanford University, California, wrote that, Surgical intervention is the only option for people with ear- should the results be replicated, the procedure represents “poten- drums that won’t heal. Tympanoplasty involves collecting graft tially the greatest advance in otology since the invention of the material from the patient to use as a patch over the perforation. -

Study Guide Medical Terminology by Thea Liza Batan About the Author
Study Guide Medical Terminology By Thea Liza Batan About the Author Thea Liza Batan earned a Master of Science in Nursing Administration in 2007 from Xavier University in Cincinnati, Ohio. She has worked as a staff nurse, nurse instructor, and level department head. She currently works as a simulation coordinator and a free- lance writer specializing in nursing and healthcare. All terms mentioned in this text that are known to be trademarks or service marks have been appropriately capitalized. Use of a term in this text shouldn’t be regarded as affecting the validity of any trademark or service mark. Copyright © 2017 by Penn Foster, Inc. All rights reserved. No part of the material protected by this copyright may be reproduced or utilized in any form or by any means, electronic or mechanical, including photocopying, recording, or by any information storage and retrieval system, without permission in writing from the copyright owner. Requests for permission to make copies of any part of the work should be mailed to Copyright Permissions, Penn Foster, 925 Oak Street, Scranton, Pennsylvania 18515. Printed in the United States of America CONTENTS INSTRUCTIONS 1 READING ASSIGNMENTS 3 LESSON 1: THE FUNDAMENTALS OF MEDICAL TERMINOLOGY 5 LESSON 2: DIAGNOSIS, INTERVENTION, AND HUMAN BODY TERMS 28 LESSON 3: MUSCULOSKELETAL, CIRCULATORY, AND RESPIRATORY SYSTEM TERMS 44 LESSON 4: DIGESTIVE, URINARY, AND REPRODUCTIVE SYSTEM TERMS 69 LESSON 5: INTEGUMENTARY, NERVOUS, AND ENDOCRINE S YSTEM TERMS 96 SELF-CHECK ANSWERS 134 © PENN FOSTER, INC. 2017 MEDICAL TERMINOLOGY PAGE III Contents INSTRUCTIONS INTRODUCTION Welcome to your course on medical terminology. You’re taking this course because you’re most likely interested in pursuing a health and science career, which entails proficiencyincommunicatingwithhealthcareprofessionalssuchasphysicians,nurses, or dentists. -
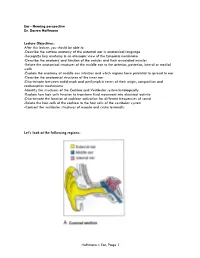
Ear, Page 1 Lecture Outline
Ear - Hearing perspective Dr. Darren Hoffmann Lecture Objectives: After this lecture, you should be able to: -Describe the surface anatomy of the external ear in anatomical language -Recognize key anatomy in an otoscopic view of the tympanic membrane -Describe the anatomy and function of the ossicles and their associated muscles -Relate the anatomical structures of the middle ear to the anterior, posterior, lateral or medial walls -Explain the anatomy of middle ear infection and which regions have potential to spread to ear -Describe the anatomical structures of the inner ear -Discriminate between endolymph and perilymph in terms of their origin, composition and reabsorption mechanisms -Identify the structures of the Cochlea and Vestibular system histologically -Explain how hair cells function to transform fluid movement into electrical activity -Discriminate the location of cochlear activation for different frequencies of sound -Relate the hair cells of the cochlea to the hair cells of the vestibular system -Contrast the vestibular structures of macula and crista terminalis Let’s look at the following regions: Hoffmann – Ear, Page 1 Lecture Outline: C1. External Ear Function: Amplification of Sound waves Parts Auricle Visible part of external ear (pinna) Helix – large outer rim Tragus – tab anterior to external auditory meatus External auditory meatus Auditory Canal/External Auditory Meatus Leads from Auricle to Tympanic membrane Starts cartilaginous, becomes bony as it enters petrous part of temporal bone Earwax (Cerumen) Complex mixture -

Vestibular Neuritis and Labyrinthitis
Vestibular Neuritis and DISORDERS Labyrinthitis: Infections of the Inner Ear By Charlotte L. Shupert, PhD with contributions from Bridget Kulick, PT and the Vestibular Disorders Association INFECTIONS Result in damage to inner ear and/or nerve. ARTICLE 079 DID THIS ARTICLE HELP YOU? SUPPORT VEDA @ VESTIBULAR.ORG Vestibular neuritis and labyrinthitis are disorders resulting from an 5018 NE 15th Ave. infection that inflames the inner ear or the nerves connecting the inner Portland, OR 97211 ear to the brain. This inflammation disrupts the transmission of sensory 1-800-837-8428 information from the ear to the brain. Vertigo, dizziness, and difficulties [email protected] with balance, vision, or hearing may result. vestibular.org Infections of the inner ear are usually viral; less commonly, the cause is bacterial. Such inner ear infections are not the same as middle ear infections, which are the type of bacterial infections common in childhood affecting the area around the eardrum. VESTIBULAR.ORG :: 079 / DISORDERS 1 INNER EAR STRUCTURE AND FUNCTION The inner ear consists of a system of fluid-filled DEFINITIONS tubes and sacs called the labyrinth. The labyrinth serves two functions: hearing and balance. Neuritis Inflamation of the nerve. The hearing function involves the cochlea, a snail- shaped tube filled with fluid and sensitive nerve Labyrinthitis Inflamation of the labyrinth. endings that transmit sound signals to the brain. Bacterial infection where The balance function involves the vestibular bacteria infect the middle organs. Fluid and hair cells in the three loop-shaped ear or the bone surrounding semicircular canals and the sac-shaped utricle and Serous the inner ear produce toxins saccule provide the brain with information about Labyrinthitis that invade the inner ear via head movement. -

Vestibular Neuritis, Labyrinthitis, and a Few Comments Regarding Sudden Sensorineural Hearing Loss Marcello Cherchi
Vestibular neuritis, labyrinthitis, and a few comments regarding sudden sensorineural hearing loss Marcello Cherchi §1: What are these diseases, how are they related, and what is their cause? §1.1: What is vestibular neuritis? Vestibular neuritis, also called vestibular neuronitis, was originally described by Margaret Ruth Dix and Charles Skinner Hallpike in 1952 (Dix and Hallpike 1952). It is currently suspected to be an inflammatory-mediated insult (damage) to the balance-related nerve (vestibular nerve) between the ear and the brain that manifests with abrupt-onset, severe dizziness that lasts days to weeks, and occasionally recurs. Although vestibular neuritis is usually regarded as a process affecting the vestibular nerve itself, damage restricted to the vestibule (balance components of the inner ear) would manifest clinically in a similar way, and might be termed “vestibulitis,” although that term is seldom applied (Izraeli, Rachmel et al. 1989). Thus, distinguishing between “vestibular neuritis” (inflammation of the vestibular nerve) and “vestibulitis” (inflammation of the balance-related components of the inner ear) would be difficult. §1.2: What is labyrinthitis? Labyrinthitis is currently suspected to be due to an inflammatory-mediated insult (damage) to both the “hearing component” (the cochlea) and the “balance component” (the semicircular canals and otolith organs) of the inner ear (labyrinth) itself. Labyrinthitis is sometimes also termed “vertigo with sudden hearing loss” (Pogson, Taylor et al. 2016, Kim, Choi et al. 2018) – and we will discuss sudden hearing loss further in a moment. Labyrinthitis usually manifests with severe dizziness (similar to vestibular neuritis) accompanied by ear symptoms on one side (typically hearing loss and tinnitus). -

Inner Ear Infection (Otitis Interna) in Dogs
Hurricane Harvey Client Education Kit Inner Ear Infection (Otitis Interna) in Dogs My dog has just been diagnosed with an inner ear infection. What is this? Inflammation of the inner ear is called otitis interna, and it is most often caused by an infection. The infectious agent is most commonly bacterial, although yeast and fungus can also be implicated in an inner ear infection. If your dog has ear mites in the external ear canal, this can ultimately cause a problem in the inner ear and pose a greater risk for a bacterial infection. Similarly, inner ear infections may develop if disease exists in one ear canal or when a benign polyp is growing from the middle ear. A foreign object, such as grass seed, may also set the stage for bacterial infection in the inner ear. Are some dogs more susceptible to inner ear infection? Dogs with long, heavy ears seem to be predisposed to chronic ear infections that ultimately lead to otitis interna. Spaniel breeds, such as the Cocker spaniel, and hound breeds, such as the bloodhound and basset hound, are the most commonly affected breeds. Regardless of breed, any dog with a chronic ear infection that is difficult to control may develop otitis interna if the eardrum (tympanic membrane) is damaged as it allows bacteria to migrate down into the inner ear. "Dogs with long, heavy ears seem to bepredisposed to chronic ear infections that ultimately lead to otitis interna." Excessively vigorous cleaning of an infected external ear canal can sometimes cause otitis interna. Some ear cleansers are irritating to the middle and inner ear and can cause signs of otitis interna if the eardrum is damaged and allows some of the solution to penetrate too deeply. -

Measuring Cochlear Duct Length – a Historical Analysis of Methods and Results Robert W
Koch et al. Journal of Otolaryngology - Head and Neck Surgery (2017) 46:19 DOI 10.1186/s40463-017-0194-2 REVIEW Open Access Measuring Cochlear Duct Length – a historical analysis of methods and results Robert W. Koch1*, Hanif M. Ladak1,2,3,4†, Mai Elfarnawany2† and Sumit K. Agrawal1,2,4,5† Abstract Background: Cochlear Duct Length (CDL) has been an important measure for the development and advancement of cochlear implants. Emerging literature has shown CDL can be used in preoperative settings to select the proper sized electrode and develop customized frequency maps. In order to improve post-operative outcomes, and develop new electrode technologies, methods of measuring CDL must be validated to allow usage in the clinic. Purpose: The purpose of this review is to assess the various techniques used to calculate CDL and provide the reader with enough information to make an informed decision on how to conduct future studies measuring the CDL. Results: The methods to measure CDL, the modality used to capture images, and the location of the measurement have all changed as technology evolved. With recent popularity and advancement in computed tomography (CT) imaging in place of histologic sections, measurements of CDL have been focused at the lateral wall (LW) instead of the organ of Corti (OC), due to the inability of CT to view intracochlear structures. After analyzing results from methods such as directly measuring CDL from histology, indirectly reconstructing the shape of the cochlea, and determining CDL based on spiral coefficients, it was determined the three dimensional (3D) reconstruction method is the most reliable method to measure CDL. -

The Eardrum Moves When the Eyes Move
bioRxiv preprint doi: https://doi.org/10.1101/156570; this version posted June 29, 2017. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The eardrum moves when the eyes move: A multisensory effect on the mechanics of hearing Short title: The eardrum moves when the eyes move K. G. Gruters*, D. L. K. Murphy*, D. W. Smith§, C. A. Shera‡, J. M. Groh*† *Department of Psychology and Neuroscience; Department of Neurobiology; Duke Institute for Brain Sciences, Duke University, Durham, NC 27708 §Program in Behavioral and Cognitive Neuroscience, Department of Psychology, University of Florida, Gainesville, FL 32611 ‡Caruso Department of Otolaryngology; Department of Physics and Astronomy, University of Southern California, Los Angeles, CA 90033 †To whom correspondence should be addressed Section: Biological Sciences: Neuroscience Acknowledgments: We are grateful to Tom Heil, Jessi Cruger, Karen Waterstradt, Christie Holmes, and Stephanie Schlebusch for technical assistance. We thank Marty Woldorff, Jeff Beck, Tobias Overath, Barbara Shinn-Cunningham, Valeria Caruso, Daniel Pages, Shawn Willett, Jeff Mohl for numerous helpful discussions and comments during the course of this project. bioRxiv preprint doi: https://doi.org/10.1101/156570; this version posted June 29, 2017. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. ABSTRACT Interactions between sensory pathways such as the visual and auditory systems are known to occur in the brain, but where they first occur is uncertain. Here we show a novel multimodal interaction evident at the eardrum. -

Tonic Tensor Tympani Syndrome (TTTS)
Tonic Tensor Tympani Syndrome (TTTS) http://www.dineenandwestcott.com.au/hyperacusis.php?fid=1 Retrieved 15ththth May 2009 In the middle ear, the tensor tympani muscle and the stapedial muscle contract to tighten the middle ear bones (the ossicles) as a reaction to loud, potentially damaging sound. This provides protection to the inner ear from these loud sounds. In many people with hyperacusis, an increased, involuntary activity can develop in the tensor tympani muscle in the middle ear as part of a protective and startle response to some sounds. This lowered reflex threshold for tensor tympani contraction is activated by the perception/anticipation of sudden, unexpected, loud sound, and is called tonic tensor tympani syndrome (TTTS). In some people with hyperacusis, it appears that the tensor tympani muscle can contract just by thinking about a loud sound. Following exposure to intolerable sounds, this heightened contraction of the tensor tympani muscle: • tightens the ear drum • stiffens the middle ear bones (ossicles) • can lead to irritability of the trigeminal nerve, which innervates the tensor tympani muscle; and to other nerves supplying the ear drum • can affect the airflow into the middle ear. The tensor tympani muscle functions in coordination with the tensor veli palatini muscle. When we yawn or swallow, these muscles work together to open the Eustachian tube. This keeps the ears healthy by clearing the middle ear of any accumulated fluid and allows the ears to “pop” by equalising pressure caused by altitude changes. TTTS can lead to a range of symptoms in and around the ear(s): ear pain; pain in the jaw joint and down the neck; a fluttering sensation in the ear; a sensation of fullness in the ear; burning/numbness/tingling in and around the ear; unsteadiness; distorted hearing.