Comparison of Acute Proton, Photon, and Low-Dose Priming Effects On
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Paxillin Binding to the Cytoplasmic Domain of CD103 Promotes Cell Adhesion and Effector
Author Manuscript Published OnlineFirst on October 11, 2017; DOI: 10.1158/0008-5472.CAN-17-1487 Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Paxillin binding to the cytoplasmic domain of CD103 promotes cell adhesion and effector functions for CD8+ resident memory T cells in tumors Ludiane Gauthier1, Stéphanie Corgnac1, Marie Boutet1, Gwendoline Gros1, Pierre Validire2, Georges Bismuth3 and Fathia Mami-Chouaib1 1 INSERM UMR 1186, Integrative Tumor Immunology and Genetic Oncology, Gustave Roussy, EPHE, Fac. de médecine - Univ. Paris-Sud, Université Paris-Saclay, 94805, Villejuif, France 2 Institut Mutualiste Montsouris, Service d’Anatomie pathologique, 75014 Paris, France. 3 INSERM U1016, CNRS UMR8104, Université Paris Descartes, Institut Cochin, 75014 Paris. S Corgnac, M Boutet and G Gros contributed equally to this work. M Boutet current address: Department of Microbiology and Immunology Albert Einstein College of Medecine, NY 10461 USA. Corresponding author: Fathia Mami-Chouaib, INSERM UMR 1186, Gustave Roussy. 39, rue Camille Desmoulins, F-94805 Villejuif. Phone: +33 1 42 11 49 65, Fax: +33 1 42 11 52 88, e-mail: [email protected] and [email protected] Running title: CD103 signaling in human TRM cells Key words: TRM cells, CD103 integrin, T-cell function and signaling, paxillin. Abbreviations: IS: immune synapse; LFA: leukocyte function-associated antigen; FI: fluorescence intensity; mAb: monoclonal antibody; phospho: phosphorylated; Pyk2: proline- rich tyrosine kinase-2; NSCLC: non-small-cell lung carcinoma; r: recombinant; sh-pxn: shorthairpin RNA-paxillin; TCR: T-cell receptor; TIL: tumor-infiltrating lymphocyte; TRM: tissue-resident memory T. -

Human Integrin Alpha L Gene Cdna Clone Plasmid
Human Integrin alpha L Gene cDNA clone plasmid Catalog Number: HG10812-M General Information Plasmid Resuspension protocol Gene : integrin, alpha L (antigen CD11A (p180), lymphocyte function-associated antigen 1; 1.Centrifuge at 5,000×g for 5 min. alpha polypeptide) 2.Carefully open the tube and add 100 l of sterile water to dissolve the DNA. Official Symbol : ITGAL 3.Close the tube and incubate for 10 minutes at room temperature. 4.Briefly vortex the tube and then do a quick spin to concentrate Synonym : CD11A, LFA-1, LFA1A, ITGAL the liquid at the bottom. Speed is less than 5000×g. 5.Store the plasmid at -20 ℃. Source : Human The plasmid is ready for: cDNA Size: 3513bp • Restriction enzyme digestion • PCR amplification RefSeq : NM_002209.2 • E. coli transformation • DNA sequencing Plasmid: pMD-ITGAL E.coli strains for transformation (recommended Description but not limited) Lot : Please refer to the label on the tube Most commercially available competent cells are appropriate for Sequence Description : the plasmid, e.g. TOP10, DH5α and TOP10F´. Identical with the Gene Bank Ref. ID sequence except for the point mutation 2928 A/G not causing the amino acid variation. Vector : pMD18-T Simple Shipping carrier : Each tube contains approximately 10 μg of lyophilized plasmid. Storage : The lyophilized plasmid can be stored at ambient temperature for three months. Quality control : The plasmid is confirmed by full-length sequencing with primers in the sequencing primer list. Sequencing primer list : M13-47 : 5’ GCCAGGGTTTTCCCAGTCACGAC 3’ RV-M : 5’ GAGCGGATAACAATTTCACACAGG 3’ Other M13 primers can also be used as sequencing primers. -

Supplementary Table 1: Adhesion Genes Data Set
Supplementary Table 1: Adhesion genes data set PROBE Entrez Gene ID Celera Gene ID Gene_Symbol Gene_Name 160832 1 hCG201364.3 A1BG alpha-1-B glycoprotein 223658 1 hCG201364.3 A1BG alpha-1-B glycoprotein 212988 102 hCG40040.3 ADAM10 ADAM metallopeptidase domain 10 133411 4185 hCG28232.2 ADAM11 ADAM metallopeptidase domain 11 110695 8038 hCG40937.4 ADAM12 ADAM metallopeptidase domain 12 (meltrin alpha) 195222 8038 hCG40937.4 ADAM12 ADAM metallopeptidase domain 12 (meltrin alpha) 165344 8751 hCG20021.3 ADAM15 ADAM metallopeptidase domain 15 (metargidin) 189065 6868 null ADAM17 ADAM metallopeptidase domain 17 (tumor necrosis factor, alpha, converting enzyme) 108119 8728 hCG15398.4 ADAM19 ADAM metallopeptidase domain 19 (meltrin beta) 117763 8748 hCG20675.3 ADAM20 ADAM metallopeptidase domain 20 126448 8747 hCG1785634.2 ADAM21 ADAM metallopeptidase domain 21 208981 8747 hCG1785634.2|hCG2042897 ADAM21 ADAM metallopeptidase domain 21 180903 53616 hCG17212.4 ADAM22 ADAM metallopeptidase domain 22 177272 8745 hCG1811623.1 ADAM23 ADAM metallopeptidase domain 23 102384 10863 hCG1818505.1 ADAM28 ADAM metallopeptidase domain 28 119968 11086 hCG1786734.2 ADAM29 ADAM metallopeptidase domain 29 205542 11085 hCG1997196.1 ADAM30 ADAM metallopeptidase domain 30 148417 80332 hCG39255.4 ADAM33 ADAM metallopeptidase domain 33 140492 8756 hCG1789002.2 ADAM7 ADAM metallopeptidase domain 7 122603 101 hCG1816947.1 ADAM8 ADAM metallopeptidase domain 8 183965 8754 hCG1996391 ADAM9 ADAM metallopeptidase domain 9 (meltrin gamma) 129974 27299 hCG15447.3 ADAMDEC1 ADAM-like, -

CD Markers Are Routinely Used for the Immunophenotyping of Cells
ptglab.com 1 CD MARKER ANTIBODIES www.ptglab.com Introduction The cluster of differentiation (abbreviated as CD) is a protocol used for the identification and investigation of cell surface molecules. So-called CD markers are routinely used for the immunophenotyping of cells. Despite this use, they are not limited to roles in the immune system and perform a variety of roles in cell differentiation, adhesion, migration, blood clotting, gamete fertilization, amino acid transport and apoptosis, among many others. As such, Proteintech’s mini catalog featuring its antibodies targeting CD markers is applicable to a wide range of research disciplines. PRODUCT FOCUS PECAM1 Platelet endothelial cell adhesion of blood vessels – making up a large portion molecule-1 (PECAM1), also known as cluster of its intracellular junctions. PECAM-1 is also CD Number of differentiation 31 (CD31), is a member of present on the surface of hematopoietic the immunoglobulin gene superfamily of cell cells and immune cells including platelets, CD31 adhesion molecules. It is highly expressed monocytes, neutrophils, natural killer cells, on the surface of the endothelium – the thin megakaryocytes and some types of T-cell. Catalog Number layer of endothelial cells lining the interior 11256-1-AP Type Rabbit Polyclonal Applications ELISA, FC, IF, IHC, IP, WB 16 Publications Immunohistochemical of paraffin-embedded Figure 1: Immunofluorescence staining human hepatocirrhosis using PECAM1, CD31 of PECAM1 (11256-1-AP), Alexa 488 goat antibody (11265-1-AP) at a dilution of 1:50 anti-rabbit (green), and smooth muscle KD/KO Validated (40x objective). alpha-actin (red), courtesy of Nicola Smart. PECAM1: Customer Testimonial Nicola Smart, a cardiovascular researcher “As you can see [the immunostaining] is and a group leader at the University of extremely clean and specific [and] displays Oxford, has said of the PECAM1 antibody strong intercellular junction expression, (11265-1-AP) that it “worked beautifully as expected for a cell adhesion molecule.” on every occasion I’ve tried it.” Proteintech thanks Dr. -

Anti-CD18 / LFA1 Beta Antibody (ARG41484)
Product datasheet [email protected] ARG41484 Package: 100 μl anti-CD18 / LFA1 beta antibody Store at: -20°C Summary Product Description Rabbit Polyclonal antibody recognizes CD18 / LFA1 beta Tested Reactivity Hu, Ms, Rat Tested Application ICC/IF, IHC-P, WB Host Rabbit Clonality Polyclonal Isotype IgG Target Name CD18 / LFA1 beta Antigen Species Human Immunogen Recombinant protein of Human CD18 / LFA1 beta. Conjugation Un-conjugated Alternate Names MF17; LAD; CD antigen CD18; MFI7; MAC-1; Cell surface adhesion glycoproteins LFA-1/CR3/p150,95 subunit beta; LCAMB; Integrin beta-2; Complement receptor C3 subunit beta; LFA-1; CD18 Application Instructions Application table Application Dilution ICC/IF 1:50 - 1:200 IHC-P 1:50 - 1:200 WB 1:500 - 1:2000 Application Note * The dilutions indicate recommended starting dilutions and the optimal dilutions or concentrations should be determined by the scientist. Positive Control Mouse thymus Calculated Mw 85 kDa Observed Size ~ 98 kDa Properties Form Liquid Purification Affinity purified. Buffer PBS (pH 7.3), 0.02% Sodium azide and 50% Glycerol. Preservative 0.02% Sodium azide Stabilizer 50% Glycerol Storage instruction For continuous use, store undiluted antibody at 2-8°C for up to a week. For long-term storage, aliquot and store at -20°C. Storage in frost free freezers is not recommended. Avoid repeated freeze/thaw www.arigobio.com 1/3 cycles. Suggest spin the vial prior to opening. The antibody solution should be gently mixed before use. Note For laboratory research only, not for drug, diagnostic or other use. Bioinformation Gene Symbol ITGB2 Gene Full Name integrin, beta 2 (complement component 3 receptor 3 and 4 subunit) Background This gene encodes an integrin beta chain, which combines with multiple different alpha chains to form different integrin heterodimers. -

Human ITGAL Peptide (DAG-P0325) This Product Is for Research Use Only and Is Not Intended for Diagnostic Use
Human ITGAL peptide (DAG-P0325) This product is for research use only and is not intended for diagnostic use. PRODUCT INFORMATION Antigen Description ITGAL encodes the integrin alpha L chain. Integrins are heterodimeric integral membrane proteins composed of an alpha chain and a beta chain. This I-domain containing alpha integrin combines with the beta 2 chain (ITGB2) to form the integrin lymphocyte function-associated antigen-1 (LFA-1), which is expressed on all leukocytes. LFA-1 plays a central role in leukocyte intercellular adhesion through interactions with its ligands, ICAMs 1-3 (intercellular adhesion molecules 1 through 3), and also functions in lymphocyte costimulatory signaling. Two transcript variants encoding different isoforms have been found for this gene. [provided by RefSeq, Jul 2008] Specificity Leukocytes. Nature Synthetic Expression System N/A Purity 70 - 90% by HPLC. Conjugate Unconjugated Sequence Similarities Belongs to the integrin alpha chain family.Contains 7 FG-GAP repeats.Contains 1 VWFA domain. Cellular Localization Membrane. Procedure None Format Liquid Preservative None Storage Shipped at 4°C. Upon delivery aliquot and store at -20°C or -80°C. Avoid repeated freeze / thaw cycles. Information available upon request. ANTIGEN GENE INFORMATION Gene Name ITGAL integrin, alpha L (antigen CD11A (p180), lymphocyte function-associated antigen 1; alpha polypeptide) [ Homo sapiens (human) ] 45-1 Ramsey Road, Shirley, NY 11967, USA Email: [email protected] Tel: 1-631-624-4882 Fax: 1-631-938-8221 1 © Creative -
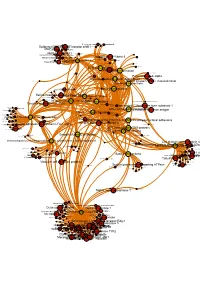
Brcaallwithlabelsintegrina1b1
Receptor-type tyrosine-proteinMuscle- skeletal receptor phosphatase tyrosine protein kinase S Dual specificity mitogen-activated protein kinase kinase 1 Plasminogen activator inhibitor-1 Epidermal growth factor receptor erbB1 Beta amyloid A4 protein MAPDual specificity Epidermalkinase mitogen-activated growth factorCasein proteinreceptor ERK2 kinase and ErbB2kinase; II (HER1 alphaMEK1/2 and (prime) HER2) Casein kinase II alpha Integrin alpha-IIb MAPCasein kinase kinase II beta ERK1 Dual specificity mitogen-activated protein kinase kinase 2 Lysyl oxidase Integrin alpha-IIb/beta-3 Furin Mitogen-activatedSerine/threonine-protein kinase RAF and Dual specificityEpidermal protein mitogen-activated growth proteinkinase; factor kinase kinase receptor 1 (Raf/MEK)ERK1/ERK2 Integrin alpha-V/beta-3 Integrin alpha-V/beta-3 and alpha-IIb/beta 3 Casein kinase II Signal transduction by L1 Integrin alpha-V/beta-6 Casein kinase II alpha/beta Vitronectin receptor alpha Bone morphogenetic protein 2 ECM proteoglycans Bone morphogenetic protein 4 VitronectinElastic fibre formation Peripheral plasma membrane protein CASK Neuropilin-1 Integrin alpha-V/beta-5 Protein kinaseProtein kinase C (PKC) alpha Integrin alpha-2/beta-3 Molecules associated with elastic fibres PKC alpha and beta-2 MER intracellular domain/EGFR extracellular domain chimera Fibronectin receptor alpha Protein kinase C- PKC; classical/novel Syndecan interactions Integrin alpha-2 von Willebrand factorIntegrin alpha-10 Laminin interactions Integrin alpha-11 Fibrinogen beta chain Serine/threonine-proteinVascular -

ICAM-1 (Phospho Tyr512) Polyclonal Antibody
ICAM-1 Monoclonal Antibody Catalog No : YM1051 Reactivity : Human Applications : WB,IF/ICC Gene Name : ICAM1 Protein Name : Intercellular adhesion molecule 1 Human Gene Id : 3383 Human Swiss Prot P05362 No : Mouse Swiss Prot P13597 No : Immunogen : Purified recombinant human ICAM-1 (N-terminus) protein fragments expressed in E.coli. Specificity : ICAM-1 Monoclonal Antibody detects endogenous levels of ICAM-1 protein. Formulation : Purified mouse monoclonal in buffer containing 0.1M Tris-Glycine (pH 7.4, 150 mM NaCl) with 0.2% sodium azide, 50% glycerol. Source : Mouse Dilution : Western Blot: 1/1000 - 1/2000. Immunofluorescence: 1/100 - 1/500. Not yet tested in other applications. Purification : Affinity purification Concentration : 1 mg/ml Storage Stability : -20°C/1 year Cell Pathway : Cell adhesion molecules (CAMs),Natural killer cell mediated cytotoxicity,Leukocyte transendothelial migration,Viral myocarditis, 1 / 2 Background : intercellular adhesion molecule 1(ICAM1) Homo sapiens This gene encodes a cell surface glycoprotein which is typically expressed on endothelial cells and cells of the immune system. It binds to integrins of type CD11a / CD18, or CD11b / CD18 and is also exploited by Rhinovirus as a receptor. [provided by RefSeq, Jul 2008], Function : function:ICAM proteins are ligands for the leukocyte adhesion protein LFA-1 (integrin alpha-L/beta-2). During leukocyte trans-endothelial migration, ICAM1 engagement promotes the assembly of endothelial apical cups through SGEF and RHOG activation. In case of rhinovirus infection acts as a cellular receptor for the virus.,online information:ICAM-1,online information:Icosahedral capsid structure,online information:Intercellular adhesion molecule entry,polymorphism:Homozygotes with ICAM1-Kalifi Met-56 seem to have an increased risk for cerebral malaria.,PTM:Monoubiquitinated, which is promoted by MARCH9 and leads to endocytosis.,similarity:Belongs to the immunoglobulin superfamily. -

CD11/CD18 (24) FITC FITC Labeled Mouse Monoclonal Antibody
Product Data Sheet IMQ-84601_2015.09-v1 CD11/CD18 (24) FITC FITC Labeled Mouse Monoclonal Antibody Catalog Number: IMQ-84601 Size: 50 µl (1 mg/ml) Class: Monoclonal Type: Antibody Clone: 24 Host / Isotype: Mouse / IgG1 Immunogen: Fibronectin-purified human monocytes Myeloma / fusion Cells from immunized Balb/c mice were fused with the Sp2/0-Ag.14 myeloma partners: cell line Species Reactivity: Human Specificity: This antibody recognises integrin alpha L,M,X/Beta 2 (CD11/CD18). Integrins are heterodimeric cell surface receptors composed of alpha and beta subunits which mediate cell-cell and cell-extracellular matrix attachments. Integrin beta2 (CD18) associates with integrin alpha L (CD11a) to form the leukocyte function- associated antigen-1 (LFA-1) with integrin alpha M (CD11b) to form Complement Receptor 3 (CR3) and with integrin alpha X (CD11c) to form Complement Receptor 4 (CR4). Each integrin receptor can bind various Intercellular adhesion molecules (ICAMs). Aberrant integrin expression has been found in many epithelial tumours. Changes in integrin expression have been shown to be important for the growth and early metastatic capacity of melanoma cells. Purification: Purified on protein A from tissue culture supernatant. Format: Purified IgG conjugated to Fluoroscein using Innova Biosciences Lightning- Link®, supplied in Phosphate buffered saline (PBS) containing 0.09% Sodium azide Applications: Flow Cytometry, Immunocytochemistry, Immunofluorescence Dilutions: Optimal antibody dilution should be determined by titration Storage: Store stock solution of the antibody at 2-8 °C in the dark. Do not freeze. References: Hogg N, et al. An anti-human monocyte/macrophage monoclonal antibody, reacting most strongly with macrophages in lymphoid tissue. -

Cx3cr1 Mediates the Development of Monocyte-Derived Dendritic Cells During Hepatic Inflammation
CX3CR1 MEDIATES THE DEVELOPMENT OF MONOCYTE-DERIVED DENDRITIC CELLS DURING HEPATIC INFLAMMATION. Supplementary material Supplementary Figure 1: Liver CD45+ myeloid cells were pre-gated for Ly6G negative cells for excluding granulocytes and HDCs subsequently analyzed among the cells that were CD11c+ and had high expression of MHCII. Supplementary Table 1 low/- high + Changes in gene expression between CX3CR1 and CX3CR1 CD11b myeloid hepatic dendritic cells (HDCs) from CCl4-treated mice high Genes up-regulated in CX3CR1 HDCs Gene Fold changes P value Full name App 4,01702 5,89E-05 amyloid beta (A4) precursor protein C1qa 9,75881 1,69E-22 complement component 1, q subcomponent, alpha polypeptide C1qb 9,19882 3,62E-20 complement component 1, q subcomponent, beta polypeptide Ccl12 2,51899 0,011769 chemokine (C-C motif) ligand 12 Ccl2 6,53486 6,37E-11 chemokine (C-C motif) ligand 2 Ccl3 4,99649 5,84E-07 chemokine (C-C motif) ligand 3 Ccl4 4,42552 9,62E-06 chemokine (C-C motif) ligand 4 Ccl6 3,9311 8,46E-05 chemokine (C-C motif) ligand 6 Ccl7 2,60184 0,009272 chemokine (C-C motif) ligand 7 Ccl9 4,17294 3,01E-05 chemokine (C-C motif) ligand 9 Ccr2 3,35195 0,000802 chemokine (C-C motif) receptor 2 Ccr5 3,23358 0,001222 chemokine (C-C motif) receptor 5 Cd14 6,13325 8,61E-10 CD14 antigen Cd36 2,94367 0,003243 CD36 antigen Cd44 4,89958 9,60E-07 CD44 antigen Cd81 6,49623 8,24E-11 CD81 antigen Cd9 3,06253 0,002195 CD9 antigen Cdkn1a 4,65279 3,27E-06 cyclin-dependent kinase inhibitor 1A (P21) Cebpb 6,6083 3,89E-11 CCAAT/enhancer binding protein (C/EBP), -

PDF Download
Integrin β2 Polyclonal Antibody Catalog No : YT2369 Reactivity : Human,Mouse,Rat Applications : IF/ICC,ELISA Gene Name : ITGB2 Protein Name : Integrin beta-2 Human Gene Id : 3689 Human Swiss Prot P05107 No : Mouse Gene Id : 16414 Mouse Swiss Prot P11835 No : Immunogen : The antiserum was produced against synthesized peptide derived from human CD18/ITGB2. AA range:720-769 Specificity : Integrin β2 Polyclonal Antibody detects endogenous levels of Integrin β2 protein. Formulation : Liquid in PBS containing 50% glycerol, 0.5% BSA and 0.02% sodium azide. Source : Rabbit Dilution : Immunofluorescence: 1/200 - 1/1000. ELISA: 1/5000. Not yet tested in other applications. Purification : The antibody was affinity-purified from rabbit antiserum by affinity- chromatography using epitope-specific immunogen. Concentration : 1 mg/ml Storage Stability : -20°C/1 year Molecularweight : 84782 1 / 2 Cell Pathway : Cell adhesion molecules (CAMs),Natural killer cell mediated cytotoxicity,Leukocyte transendothelial migration,Regulates Actin and Cytoskeleton,Viral myocarditis, Background : integrin subunit beta 2(ITGB2) Homo sapiens This gene encodes an integrin beta chain, which combines with multiple different alpha chains to form different integrin heterodimers. Integrins are integral cell-surface proteins that participate in cell adhesion as well as cell-surface mediated signalling. The encoded protein plays an important role in immune response and defects in this gene cause leukocyte adhesion deficiency. Alternative splicing results in multiple transcript variants. [provided by RefSeq, Dec 2014], Function : disease:Defects in ITGB2 are the cause of leukocyte adhesion deficiency type I (LAD1) [MIM:116920]. LAD1 patients have recurrent bacterial infections and their leukocytes are deficient in a wide range of adhesion-dependent functions.,function:Integrin alpha-L/beta-2 is a receptor for ICAM1, ICAM2, ICAM3 and ICAM4. -

Anti-CD54 / ICAM1 Antibody (ARG40213)
Product datasheet [email protected] ARG40213 Package: 100 μl anti-CD54 / ICAM1 antibody Store at: -20°C Summary Product Description Rabbit Polyclonal antibody recognizes CD54 / ICAM1 Tested Reactivity Hu, Ms, Rat Tested Application ICC/IF, WB Host Rabbit Clonality Polyclonal Isotype IgG Target Name CD54 / ICAM1 Antigen Species Human Immunogen Recombinant protein of Human CD54 / ICAM1. (NP_000192.2) Conjugation Un-conjugated Alternate Names CD54; CD antigen CD54; BB2; P3.58; Intercellular adhesion molecule 1; Major group rhinovirus receptor; ICAM-1 Application Instructions Application table Application Dilution ICC/IF 1:50 - 1:200 WB 1:500 - 1:2000 Application Note * The dilutions indicate recommended starting dilutions and the optimal dilutions or concentrations should be determined by the scientist. Positive Control Rat lung, Mouse lung and Raji Calculated Mw 58 kDa Properties Form Liquid Purification Affinity purified. Buffer PBS (pH 7.3), 0.02% Sodium azide and 50% Glycerol. Preservative 0.02% Sodium azide Stabilizer 50% Glycerol Storage instruction For continuous use, store undiluted antibody at 2-8°C for up to a week. For long-term storage, aliquot and store at -20°C. Storage in frost free freezers is not recommended. Avoid repeated freeze/thaw cycles. Suggest spin the vial prior to opening. The antibody solution should be gently mixed before use. Note For laboratory research only, not for drug, diagnostic or other use. www.arigobio.com 1/2 Bioinformation Gene Symbol ICAM1 Gene Full Name intercellular adhesion molecule 1 Background This gene encodes a cell surface glycoprotein which is typically expressed on endothelial cells and cells of the immune system.