Influences of Flood and Ebb Tides on Nutrient Fluxes and Chlorophyll on an Intertidal Flat
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Nahant Reconnaissance Report
NAHANT RECONNAISSANCE REPORT ESSEX COUNTY LANDSCAPE INVENTORY MASSACHUSETTS HERITAGE LANDSCAPE INVENTORY PROGRAM Massachusetts Department of Conservation and Recreation Essex National Heritage Commission PROJECT TEAM Massachusetts Department of Conservation and Recreation Jessica Rowcroft, Preservation Planner Division of Planning and Engineering Essex National Heritage Commission Bill Steelman, Director of Heritage Preservation Project Consultants Shary Page Berg Gretchen G. Schuler Virginia Adams, PAL Local Project Coordinator Linda Pivacek Local Heritage Landscape Participants Debbie Aliff John Benson Mark Cullinan Dan deStefano Priscilla Fitch Jonathan Gilman Tom LeBlanc Michael Manning Bill Pivacek Linda Pivacek Emily Potts Octavia Randolph Edith Richardson Calantha Sears Lynne Spencer Julie Stoller Robert Wilson Bernard Yadoff May 2005 INTRODUCTION Essex County is known for its unusually rich and varied landscapes, which are represented in each of its 34 municipalities. Heritage landscapes are those places that are created by human interaction with the natural environment. They are dynamic and evolving; they reflect the history of the community and provide a sense of place; they show the natural ecology that influenced the land use in a community; and heritage landscapes often have scenic qualities. This wealth of landscapes is central to each community’s character; yet heritage landscapes are vulnerable and ever changing. For this reason it is important to take the first steps toward their preservation by identifying those landscapes that are particularly valued by the community – a favorite local farm, a distinctive neighborhood or mill village, a unique natural feature, an inland river corridor or the rocky coast. To this end, the Massachusetts Department of Conservation and Recreation (DCR) and the Essex National Heritage Commission (ENHC) have collaborated to bring the Heritage Landscape Inventory program (HLI) to communities in Essex County. -

NEERS 2017 Spring Meeting March 16 – 18, 2017 University of Connecticut, Avery Point Campus Groton, CT
NEERS 2017 Spring Meeting March 16 – 18, 2017 University of Connecticut, Avery Point Campus Groton, CT Organized and Hosted By: Jamie Vaudrey, University of Connecticut Sarah Crosby, Harbor Watch Craig Tobias & Michael Whitney of UCONN ABSTRACTS Alldred*, M. (1), Hoellein, T. (2), Bruesewitz, D. (3), and Zarnoch, C. (1); (1) Baruch College, City University of New York, New York, NY; (2) Loyola University, Chicago, IL; (3) Colby College, Waterville, ME. [email protected] NITROGEN-REMOVAL SERVICES OF RESTORED SALT MARSHES IN JAMAICA BAY (NEW YORK, NY) Coastal wetlands are important sites of nitrogen removal, a critical ecosystem service in highly eutrophic environments. In Jamaica Bay, over 92% of historic wetland area has been lost over the past century. Despite considerable efforts to restore wetland ecosystems in Jamaica Bay and throughout New York City, few studies have examined the value of ecosystem services used to justify their cost, and little is known about the ecological mechanisms contributing to the success or failure of reconstruction. Past and ongoing restoration efforts in Jamaica Bay provide a unique opportunity to study nitrogen-removal ecosystem services in natural and restored wetlands in an urban, eutrophic environment. In collaboration with researchers from several institutions, we are using a chronosequence of marsh restorations to assess how marsh vegetation, sediment characteristics, and key processes of the nitrogen cycle develop over time following restoration. The goal of our project is to determine the restoration age and environmental conditions under which salt-marsh restoration will effectively provide ecosystem services such as nitrogen removal. We employ a combination of flow-through incubation, field survey, and experimental methods to identify the key biological and abiotic factors limiting nitrogen-removal services in natural and restored marshes. -

Massachusetts Ocean Management Plan
Massachusetts Ocean Management Plan Volume 2 Baseline Assessment and Science Framework December 2009 Introduction Volume 2 of the Massachusetts Ocean Management Plan focuses on the data and scientific aspects of the plan and its implementation. It includes these two separate documents: • Baseline Assessment of the Massachusetts Ocean Planning Area - This Oceans Act-mandated product includes information cataloging the current state of knowledge regarding human uses, natural resources, and other ecosystem factors in Massachusetts ocean waters. • Science Framework - This document provides a blueprint for ocean management- related science and research needs in Massachusetts, including priorities for the next five years. i Baseline Assessment of the Massachusetts Ocean Management Planning Area Acknowledgements The authors thank Emily Chambliss and Dan Sampson for their help in preparing Geographic Information System (GIS) data for presentation in the figures. We also thank Anne Donovan and Arden Miller, who helped with the editing and layout of this document. Special thanks go to Walter Barnhardt, Ed Bell, Michael Bothner, Erin Burke, Tay Evans, Deb Hadden, Dave Janik, Matt Liebman, Victor Mastone, Adrienne Pappal, Mark Rousseau, Tom Shields, Jan Smith, Page Valentine, John Weber, and Brad Wellock, who helped us write specific sections of this assessment. We are grateful to Wendy Leo, Peter Ralston, and Andrea Rex of the Massachusetts Water Resources Authority for data and assistance writing the water quality subchapter. Robert Buchsbaum, Becky Harris, Simon Perkins, and Wayne Petersen from Massachusetts Audubon provided expert advice on the avifauna subchapter. Kevin Brander, David Burns, and Kathleen Keohane from the Massachusetts Department of Environmental Protection and Robin Pearlman from the U.S. -

Northeast Region Bacteria Source Tracking 2007 Results
DRAFT Technical Memorandum Northeast Region Bacteria Source Tracking 2007 Results DWM Control Number: CN XXX.X Prepared By: Jenny Birnbaum Katie Zink January 2008 Commonwealth of Massachusetts Executive Office of Environmental Affairs Ian A. Bowles, Secretary Massachusetts Department Of Environmental Protection Laurie Burt, Commissioner Bureau of Resource Protection Glenn Haas, Acting Assistant Commissioner Division of Watershed Management Glenn Haas, Director Table of Contents EXECUTIVE SUMMARY ................................................................................................................... 6 ACKNOWLEDGEMENTS................................................................................................................... 9 1.0 INTRODUCTION ....................................................................................................................... 10 2.0 METHODS................................................................................................................................12 3.0 SITE DESCRIPTIONS, RESULTS, AND SIGNIFICANT FINDINGS ............................................... 13 3.1 Boston Harbor - Mystic..........................................................................................................................13 Mill Brook (Arlington) ......................................................................................................... 13 Sub-watershed Description ............................................................................................... 13 Results.............................................................................................................................. -

Mystic River Watershed with Myrwa
Silver Lake DANVERS Crane River Ipswich River Crystal Lake Elginwood Pond Waters River 95 38 Devils Dishfull Pond Patches Pond Winona Pond PEABODY LYNNFIELD Puritan Lawn Pond WILMINGTON Pierces Pond 128 Pillings Pond 28 Suntaug Lake Colonial GC Ponds Cedar Pond Proctor Pond Goldthwait Brook Baltic Pond READING Stoneys Pond Lake Q uannapow itt Phillips Pond Bartholomew Pond Spring Pond Mill Pond Reservoir Mishawum Lake Hawkes Brook Pond Browns Pond BURLINGTON Hawkes Pond Cedar Pond SALEM 95 Spring Pond Lily Mere Pond WAKEFIELD Wa lde n Pond Sluice Pond Crystal Lake ABR049 Pearce Lake Camo Nihan Pond Floating Bridge Pond Silver Lake Flax Pond WOBURN Whittemore Pond Spring Pond Breeds Pond LYNN Birch Pond Griswold Pond Prankers Pond 93 STONEHAM Buckman Pond SAUGUS Doleful Pond Butterfields Pond Horn Pond Stevens Pond ABR028 Shute Pond North Reservoir Ell Pond Saugus River Dark Hollow Pond Long Pond Aberjona River Spot Pond Middle Reservoir Winter Pond MELROSE Long Pond Judkins Pond 2nd Pond 1st Pond 3rd Pond Wedge Pond Towners Pond L y n n H a r b o r Bear Creek WINCHESTER Swains Pond Quarter Mile Pond Fells Reservoir Nahant Bay ABR006 South Reservoir Wrights Pond Pines River Pines River LEXINGTON Upper Pines Pond Mystic Lake Diamond Creek Brooks Pond MALDEN UPL001 38 Fellsmere Pond Pines River 2A NAHANT Arlington Reservoir Lower Mystic Lake MEDFORD MIB001 REVERE MYR071 MAR036 Broad Sound ARLINGTON MEB001 60 2 1 Malden River Spy Pond EVERETT MIC004 3 ALB006 Mill Creek Chelsea River Little Pond 28 Mystic River WIB001 Alew ife Brook CHELSEA -
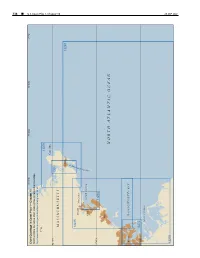
CPB1 C10 WEB.Pdf
338 ¢ U.S. Coast Pilot 1, Chapter 10 Chapter 1, Pilot Coast U.S. 70°45'W 70°30'W 70°15'W 71°W Chart Coverage in Coast Pilot 1—Chapter 10 NOAA’s Online Interactive Chart Catalog has complete chart coverage http://www.charts.noaa.gov/InteractiveCatalog/nrnc.shtml 71°W 13279 Cape Ann 42°40'N 13281 MASSACHUSETTS Gloucester 13267 R O B R A 13275 H Beverly R Manchester E T S E C SALEM SOUND U O Salem L G 42°30'N 13276 Lynn NORTH ATLANTIC OCEAN Boston MASSACHUSETTS BAY 42°20'N 13272 BOSTON HARBOR 26 SEP2021 13270 26 SEP 2021 U.S. Coast Pilot 1, Chapter 10 ¢ 339 Cape Ann to Boston Harbor, Massachusetts (1) This chapter describes the Massachusetts coast along and 234 miles from New York. The entrance is marked on the northwestern shore of Massachusetts Bay from Cape its eastern side by Eastern Point Light. There is an outer Ann southwestward to but not including Boston Harbor. and inner harbor, the former having depths generally of The harbors of Gloucester, Manchester, Beverly, Salem, 18 to 52 feet and the latter, depths of 15 to 24 feet. Marblehead, Swampscott and Lynn are discussed as are (11) Gloucester Inner Harbor limits begin at a line most of the islands and dangers off the entrances to these between Black Rock Danger Daybeacon and Fort Point. harbors. (12) Gloucester is a city of great historical interest, the (2) first permanent settlement having been established in COLREGS Demarcation Lines 1623. The city limits cover the greater part of Cape Ann (3) The lines established for this part of the coast are and part of the mainland as far west as Magnolia Harbor. -

AUGUST 2007 • Page 1 Nahant Harbor Review
NAHANT HARBOR REVIEW • AUGUST 2007 • Page 1 Nahant Harbor Review A monthly publication, in service since March 1994, dedicated to strengthening the spirit of community by serving the interests of civic, religious and business organizations of Nahant, Massachusetts, USA. Volume 14 Issue 8 AUGUST 2007 Nahanter Rachel Tarmy Celebrate 25 Years with My Brother’s Competes for Miss Teen Boston Table at Summer Garden Party Aug. 9th Rachel Tarmy of Nahant, daughter of Les and Julie Tarmy, was recently selected, to participate in The Summer Garden Party, a critical fund-raiser to benefit the North Shore’s largest Nationals’ 2007 Miss Teen Boston pageant competi- soup kitchen, My Brother’s Table, in Lynn, will take place on August 9th, at Marian tion, that will take place on July 29th, 2007. Rachel Court College, Little’s Point Road, in learned of her acceptance into this year’s competition, Swampscott, beginning at 7:00 p.m. as Nationals, Inc. Tickets start at $65. This vital event announced their includes the auction of an Italian Villa selections on Vacation, from the Parker Company, as Monday, July 2nd. well as a catered event for 50, from Rachel submitted Tiger Lily Caterers of Beverly. Please an application and RSVP Mary at 781-595-3224. took part in an This year, My Brother’s Table is interview session, celebrating 25 years of operation; it has that was con- served over two million meals to the ducted by Patty hungry, since 1982. Last year, over Niedert, this year’s 80,000 meals were served to those in Boston Pageant need. -

Provides This File for Download from Its Web Site for the Convenience of Users Only
Disclaimer The Massachusetts Department of Environmental Protection (MassDEP) provides this file for download from its Web site for the convenience of users only. Please be aware that the OFFICIAL versions of all state statutes and regulations (and many of the MassDEP policies) are only available through the State Bookstore or from the Secretary of State’s Code of Massachusetts Regulations (CMR) Subscription Service. When downloading regulations and policies from the MassDEP Web site, the copy you receive may be different from the official version for a number of reasons, including but not limited to: • The download may have gone wrong and you may have lost important information. • The document may not print well given your specific software/ hardware setup. • If you translate our documents to another word processing program, it may miss/skip/lose important information. • The file on this Web site may be out-of-date (as hard as we try to keep everything current). If you must know that the version you have is correct and up-to-date, then purchase the document through the state bookstore, the subscription service, and/or contact the appropriate MassDEP program. 314 CMR: DIVISION OF WATER POLLUTION CONTROL 4.06: continued FIGURE LIST OF FIGURES A River Basins and Coastal Drainage Areas 1 Hudson River Basin (formerly Hoosic, Kinderhook and Bashbish River Basins) 2 Housatonic River Basin 3 Farmington River Basin 4 Westfield River Basin 5 Deerfield River Basin 6 Connecticut River Basin 7 Millers River Basin 8 Chicopee River Basin 9 Quinebaug -

An Annotated List of the Fishes of Massachusetts Bay
NOAA Technical Memorandum NMFS-F/NEC-51 This TM series is used for documentation and timely communication of preliminary results, interim reports, or special purpose Information, and has not received complete formal review, editorial control, or detailed editing. An Annotated List of the Fishes of Massachusetts Bay Bruce B. Collette 1,2and Karsten E. Hartel 3 1Marine Science Center, Northeastern University, Nahant, MA 07907 2National Systematics Labbratory; National Marine Fisheries Service, National Museum 3 of Natural History, Washington, DC 20560 Museum of Comparative Zoology, Harvard University, Cambridge, MA 02 138 U.S. DEPARTMENT OF COMMERCE C. William Verity, Secretary National Oceanic and Atmospheric Administration J. Curtis Mack II, Assistant Secretary for Oceans and Atmosphere National Marine Fisheries Service William E. Evans, Assistant Administrator for Fisheries Northeast Fisheries Center Woods Hole, Massachusetts February 1988 THIS PAGE INTENTIONALLY LEFT BLANK ABSTRACT The list includes 141 species in 68 families based on authoritative literature reports and museum specimens. First records for Massachusetts Bay are recorded for: Atlantic angel shark, Squatina dumerill smooth skate, Raja senta;= wolf eelpout, Lycenchelys verrillii; lined seahorse, DHippo-us erectus; rough scad, Trachurus lathami smallmouth flounder, iii THIS PAGE INTENTIONALLY LEFT BLANK CONTENTS INTRODUCTION.................................................. 1 ANNOTATED LIST................................................ 5 Hagfishes. Family Myxinidae 1. Atlantic hagfish. Myxine slutinosa Linnaeus. 5 Lampreys. Family Petromyzontidae 2. Sea lamprey. Petromyzon marinus Linnaeus. 5 Sand sharks.Family Odontaspididae 3. Sand tiger. Euqomphodus taurus (Rafinesque) . 5 Thresher sharks. Family Alopiidae 4. Thresher shark. Alopias vulpinus (Bonnaterre) . 6 Mackerel sharks. Family Lamnidae 5. White shark. Carcharodon carcharim (Linnaeus) . 6 6. Basking shark. Cetohinus maximus (Gunnerus) . 7. Shortfin mako. -

Open Space & Recreation Plan 2020-2026
OPEN SPACE & RECREATION PLAN 2020-2026 Town of Swampscott DRAFT FOR PUBLIC COMMENT APPROVALS TOWN OF SWAMPSCOTT - TBD COMMONWEALTH OF MASSACHUSETTS - TBD Submitted on: DATE Submitted to: Executive Office of Energy and Environmental Affairs Submitted by: Swampscott Open Space & Recreation Plan Committee TABLE OF CONTENTS I. Plan Summary 7 II. Introduction 10 Statement of Purpose 10 Planning Process and Public Participation 10 III. Community Setting 13 Regional Context 13 History of the Community 15 Population Characteristics & Demographics 18 Growth and Development 22 Infrastructure 23 IV. Environmental Inventory and Analysis 29 Geology, Soils and Topography 29 Landscape Character 35 Water Resources 35 Vegetation 46 Fisheries and Wildlife 50 Scenic Resources and Unique Environments 52 Environmental Challenges 55 V. Inventory of Lands of Conservation and Recreation Interest 57 PUBLICLY OWNED AREAS 61 Beaches 61 Conservation Areas 63 Parks & Other Open Space Areas 64 Recreation Areas 67 Right-of-Ways 69 PRIVATELY-OWNED AREAS 70 Beaches 70 Conservation Areas 71 Parks & Other Open Space Areas 71 Recreation Areas/61B Land 73 Private Lands with Future Public Benefits 73 ADA Access 74 Lands of Conservation and Recreation Interest - Matrix 75 VI. Community Goals 81 Description of Process 81 Statement of Open Space & Recreation Goals 82 VII. Analysis of Needs 85 Summary of Resource Protection Needs 85 Summary of Community Needs 86 Management and Maintenance Needs 87 VIII. Goals & Objectives 90 IX. Five-Year Action Plan 94 X. Public Comments 104 -

Appendix 20 North Shore Coastal Drainage Area Assessment and Listing Decision Summary
2018-20ILApp20_NCoastal_DRAFT210323.docx Appendix 20 North Shore Coastal Drainage Area Assessment and Listing Decision Summary Massachusetts Integrated List of Waters for the Clean Water Act 2018/20 Reporting Cycle Draft for Public Comment Prepared by: Massachusetts Department of Environmental Protection Division of Watershed Management Watershed Planning Program 2018-20ILApp20_NCoastal_DRAFT210323.docx Table of Contents 2018/20 Cycle Impairment Changes ............................................................................................................. 5 Alewife Brook (MA93-26) ............................................................................................................................. 7 Alewife Brook (MA93-45) ............................................................................................................................. 8 Alewife Brook (MA93-46) ........................................................................................................................... 10 Annisquam River (MA93-12) ....................................................................................................................... 11 Babson Reservoir (MA93001) ..................................................................................................................... 12 Bass River (MA93-07) .................................................................................................................................. 13 Bass River (MA93-08) ................................................................................................................................. -

Beaches We Can Be Proud OF” a Report of the Metropolitan Beaches Commission Letter from the Chairs
Contents METROPOLITAN BEACHES COMMISSION Senator Jack Hart Letter from the Chairs 1 Co-Chairman 1. The Challenge 5 Representative Anthony Petruccelli Co-Chairman 2. The Commission’s Work 6 Kip Becker Boston University 3. Findings and Recommendations 23 Mark Cullinan 4. Conclusion 37 Nahant Town Manager Rosanne Foley List of Appendices 38 DotWell Paul Grogan The Boston Foundation Douglas Gutro Quincy City Council President Carol Haney Revere Beach Partnership Senator Robert Hedlund Plymouth & Norfolk Representative Bradford Hill Fourth Essex Salvatore LaMattina Boston City Councilor Anthony Lozzi City of Lynn Senator Thomas McGee Third Essex & Middlesex Joan Meschino Hull Board of Selectmen Secretary Daniel O’Connell Executive Office of Housing and Economic Development Joseph Orfant Department of Conservation and Recreation Representative Kathi-Anne Reinstein Sixteenth Suffolk Richard White Winthrop Town Manager “BEACHES WE CAN BE PROUD OF” A Report of the Metropolitan Beaches Commission Letter from the Chairs Dear Friends: Like many of you, the two of us grew up on these beaches. Looking back, some of our fond- est memories are of the times we spent each summer with our families and friends at Carson Beach in South Boston and Constitution Beach in East Boston. For us, like so many families across the region, the public beaches from Nantasket to Nahant were our “Cape Cod.” We spent our summer days from sunup to sundown on these beaches. We learned to swim in the ocean, and to sail at the youth sailing centers nearby. We grew to appreciate the quality time we were able to spend with our families at the beach, and the value of teamwork and community on the nearby ballfields and skating rinks.