Treatment of Particular Topics. Thus Markert Illustrates Some of the Done
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Biology Incomplete Dominance 5E Model Lesson
BIOLOGY INCOMPLETE DOMINANCE 5E MODEL LESSON Teacher: Expected Length of Lesson Topic: Unit: Heather Lesson: Incomplete Genetics Yarbrough 1 - 90 minute class Dominance SB3. Obtain, evaluate, and communicate information to analyze how biological traits are passed on to successive generations. Targeted Content Standards/ b. Use mathematical models to predict and explain patterns of inheritance. Element: (Clarification statement: Students should be able to use Punnett squares (Include the entire standard) (monohybrid and dihybrid crosses) and/or rules of probability, to analyze the following inheritance patterns: dominance, codominance, incomplete dominance.) L9-10RST2: Determine the central ideas or conclusions of a text; trace the text’s explanation or depiction of a complex process, phenomenon, or concept; provide an accurate summary of the text. L9-10RST7: Translate quantitative or technical information expressed in words in a text into visual form (e.g., a table or chart) and translate information expressed visually or mathematically (e.g., in an equation) into words. L9-10RST9: Compare and contrast findings presented in a text to those Targeted Literacy Skills or Standards: (include as many as your from other sources (including their own experiments), noting when the lesson incorporates) findings support or contradict previous explanations or accounts. L9-10WHST2: Write informative/explanatory texts, including the narration of historical events, scientific procedures/ experiments, or technical processes. d. Use precise language and domain-specific vocabulary to manage the complexity of the topic and convey a style appropriate to the discipline and context as well as to the expertise of likely readers. L9-10WHST4: Produce clear and coherent writing in which the development, organization, and style are appropriate to task, purpose, and audience. -

Genetics in Harry Potter's World: Lesson 2
Genetics in Harry Potter’s World Lesson 2 • Beyond Mendelian Inheritance • Genetics of Magical Ability 1 Rules of Inheritance • Some traits follow the simple rules of Mendelian inheritance of dominant and recessive genes. • Complex traits follow different patterns of inheritance that may involve multiples genes and other factors. For example, – Incomplete or blended dominance – Codominance – Multiple alleles – Regulatory genes Any guesses on what these terms may mean? 2 Incomplete Dominance • Incomplete dominance results in a phenotype that is a blend of a heterozygous allele pair. Ex., Red flower + Blue flower => Purple flower • If the dragons in Harry Potter have fire-power alleles F (strong fire) and F’ (no fire) that follow incomplete dominance, what are the phenotypes for the following dragon-fire genotypes? – FF – FF’ – F’F’ 3 Incomplete Dominance • Incomplete dominance results in a phenotype that is a blend of the two traits in an allele pair. Ex., Red flower + Blue flower => Purple flower • If the Dragons in Harry Potter have fire-power alleles F (strong fire) and F’ (no fire) that follow incomplete dominance, what are the phenotypes for the following dragon-fire genotypes: Genotypes Phenotypes FF strong fire FF’ moderate fire (blended trait) F’F’ no fire 4 Codominance • Codominance results in a phenotype that shows both traits of an allele pair. Ex., Red flower + White flower => Red & White spotted flower • If merpeople have tail color alleles B (blue) and G (green) that follow the codominance inheritance rule, what are possible genotypes and phenotypes? Genotypes Phenotypes 5 Codominance • Codominance results in a phenotype that shows both traits of an allele pair. -

Roadmap • Optimal Mutation Rate • Dominance and Its Implications
Roadmap • Optimal mutation rate • Dominance and its implications { Why is an allele dominant or recessive? { Overdominance (heterozygote advantage) { Underdominance (heterozygote inferiority) One minute responses • Q: I don't understand degrees of freedom! (About six of these....) • Q: Show the calculation of µ and ν Degrees of freedom revisited Thanks to Patrick Runkel: http://blog.minitab.com/blog/statistics-and-quality-data-analysis/ what-are-degrees-of-freedom-in-statistics A a Total A 20 a 10 Total 15 15 One more look at degrees of freedom Fictional data for sickle-cell hemoglobin (alleles A and S) in African-American adults Normal AA 400 Carrier AS 90 Affected SS 10 • Suppose I told you: { How many people I sampled { How many of each allele I found { How many AS carriers I found • Are there any possible surprises left in the data? (AA? SS?) • This is why there is only 1 df Mu and nu • µ (mu, forward mutation rate) { mutation rate per site is observed { rate per significant site in gene is: { rate per site x number of significant sites • ν (nu, back mutation rate) { mutation rate per site is observed { need the right nucleotide (1/3 chance) { rate per site x 1/3 Is mutation good or bad? • Most mutations have no fitness effect • Of those that do, most are bad • Most organisms expend significant energy trying to avoid mutations (DNA proofreading, etc) • Are organisms trying (and failing) to reach a mutation rate of zero? • Could there be selection in favor of a non-zero rate? Transposons as mutagens • Transposons are genetic elements that -

Directional Selection and Variation in Finite Populations
Copyright 0 1987 by the Genetics Society of America Directional Selection and Variation in Finite Populations Peter D. Keightley and ‘William G. Hill Institute of Animal Genetics, University of Edinburgh, West Mains Road, Edinburgh EH9 3JN, Scotland Manuscript received April 12, 1987 Revised copy accepted July 16, 1987 ABSTRACT Predictions are made of the equilibrium genetic variances and responses in a metric trait under the joint effects of directional selection, mutation and linkage in a finite population. The “infinitesimal model“ is analyzed as the limiting case of many mutants of very small effect, otherwise Monte Carlo simulation is used. If the effects of mutant genes on the trait are symmetrically distributed and they are unlinked, the variance of mutant effects is not an important parameter. If the distribution is skewed, unless effects or the population size is small, the proportion of mutants that have increasing effect is the critical parameter. With linkage the distribution of genotypic values in the population becomes skewed downward and the equilibrium genetic variance and response are smaller as disequilibrium becomes important. Linkage effects are greater when the mutational variance is contributed by many genes of small effect than few of large effect, and are greater when the majority of mutants increase rather than decrease the trait because genes that are of large effect or are deleterious do not segregate for long. The most likely conditions for ‘‘Muller’s ratchet” are investi- gated. N recent years there has been much interest in the is attained more quickly in the presence of selection I production and maintenance of variation in pop- than in its absence, and is highly dependent on popu- ulations by mutation, stimulated by the presence of lation size. -

Dominance Hierarchy Arising from the Evolution of a Complex Small RNA Regulatory Network Eléonore Durand, Raphaël Méheust, Marion Soucaze, Pauline M
RESEARCH ◥ between S alleles (6). Selection is expected to RESEARCH ARTICLE favor genetic elements (“dominance modifiers”), which establish dominance-recessivity interac- tion rather than codominance, because individ- PLANT GENETICS uals with a codominant genotype can produce pollen rejected by more potential mates than would occur in a dominant-recessive system (7, 8). Dominance hierarchy arising from On the basis of modeling (8), the large non- recombining region composing the S locus (9–11) is a strong candidate region for hosting such ge- the evolution of a complex small netic elements. Until recently, the dominance modifiers as- RNA regulatory network sumed in models (7, 8) remained hypothetical. A particular small RNA (sRNA) has been identified Eléonore Durand,1,2* Raphaël Méheust,1* Marion Soucaze,1 Pauline M. Goubet,1† (12) within the S locus of dominant alleles in Sophie Gallina,1 Céline Poux,1 Isabelle Fobis-Loisy,2 Eline Guillon,2 Thierry Gaude,2 Brassica (called Smi). This sRNA acts as a trans- Alexis Sarazin,3 Martin Figeac,4 Elisa Prat,5 William Marande,5 Hélène Bergès,5 modifier of the gene controlling pollen specificity Xavier Vekemans,1 Sylvain Billiard,1 Vincent Castric1‡ via de novo methylation of the promoter of re- cessive alleles, which leads to transcriptional si- The prevention of fertilization through self-pollination (or pollination by a close relative) lencing of recessive alleles by dominant alleles in the Brassicaceae plant family is determined by the genotype of the plant at the (13, 14). However, the mechanism in the more self-incompatibility locus (S locus). The many alleles at this locus exhibit a dominance complex dominance-recessivity networks in spe- Downloaded from hierarchy that determines which of the two allelic specificities of a heterozygous genotype cies that have many levels in the dominance is expressed at the phenotypic level. -

Patterns and Power of Phenotypic Selection in Nature
Articles Patterns and Power of Phenotypic Selection in Nature JOEL G. KINGSOLVER AND DAVID W. PFENNIG Phenotypic selection occurs when individuals with certain characteristics produce more surviving offspring than individuals with other characteristics. Although selection is regarded as the chief engine of evolutionary change, scientists have only recently begun to measure its action in the wild. These studies raise numerous questions: How strong is selection, and do different types of traits experience different patterns of selection? Is selection on traits that affect mating success as strong as selection on traits that affect survival? Does selection tend to favor larger body size, and, if so, what are its consequences? We explore these questions and discuss the pitfalls and future prospects of measuring selection in natural populations. Keywords: adaptive landscape, Cope’s rule, natural selection, rapid evolution, sexual selection henotypic selection occurs when individuals with selection on traits that affect survival stronger than on those Pdifferent characteristics (i.e., different phenotypes) that affect only mating success? In this article, we explore these differ in their survival, fecundity, or mating success. The idea and other questions about the patterns and power of phe- of phenotypic selection traces back to Darwin and Wallace notypic selection in nature. (1858), and selection is widely accepted as the primary cause of adaptive evolution within natural populations.Yet Darwin What is selection, and how does it work? never attempted to measure selection in nature, and in the Selection is the nonrandom differential survival or repro- century following the publication of On the Origin of Species duction of phenotypically different individuals. -

Selection and Gene Duplication: a View from the Genome Comment Andreas Wagner
http://genomebiology.com/2002/3/5/reviews/1012.1 Minireview Selection and gene duplication: a view from the genome comment Andreas Wagner Address: Department of Biology, University of New Mexico, 167A Castetter Hall, Albuquerque, NM 817131-1091, USA. E-mail: [email protected] Published: 15 April 2002 Genome Biology 2002, 3(5):reviews1012.1–1012.3 reviews The electronic version of this article is the complete one and can be found online at http://genomebiology.com/2002/3/5/reviews/1012 © BioMed Central Ltd (Print ISSN 1465-6906; Online ISSN 1465-6914) Abstract reports Immediately after a gene duplication event, the duplicate genes have redundant functions. Is natural selection therefore completely relaxed after duplication? Does one gene evolve more rapidly than the other? Several recent genome-wide studies have suggested that duplicate genes are always under purifying selection and do not always evolve at the same rate. deposited research When a gene duplication event occurs, the duplicate genes synonymous (silent) nucleotide substitutions, Ks, and have redundant functions. Many deleterious mutations may second, of non-synonymous nucleotide substitutions (which then be harmless, because even if one gene suffers a muta- change the encoded amino acid), Ka (see Box 1). The ratio tion, the redundant gene copy can provide a back-up func- Ka/Ks provides a measure of the selection pressure to which tion. Put differently, after gene duplication - which can arise a gene pair is subject. If a duplicate gene pair shows a Ka/Ks through polyploidization (whole-genome duplication), non- ratio of about 1, that is, if amino-acid replacement substitu- homologous recombination, or through the action of retro- tions occur at the same rate as synonymous substitutions, transposons - one or both duplicates should experience then few or no amino-acid replacement substitutions have refereed research relaxed selective constraints that result in elevated rates of been eliminated since the gene duplication. -

An Introduction to Quantitative Genetics I Heather a Lawson Advanced Genetics Spring2018 Outline
An Introduction to Quantitative Genetics I Heather A Lawson Advanced Genetics Spring2018 Outline • What is Quantitative Genetics? • Genotypic Values and Genetic Effects • Heritability • Linkage Disequilibrium and Genome-Wide Association Quantitative Genetics • The theory of the statistical relationship between genotypic variation and phenotypic variation. 1. What is the cause of phenotypic variation in natural populations? 2. What is the genetic architecture and molecular basis of phenotypic variation in natural populations? • Genotype • The genetic constitution of an organism or cell; also refers to the specific set of alleles inherited at a locus • Phenotype • Any measureable characteristic of an individual, such as height, arm length, test score, hair color, disease status, migration of proteins or DNA in a gel, etc. Nature Versus Nurture • Is a phenotype the result of genes or the environment? • False dichotomy • If NATURE: my genes made me do it! • If NURTURE: my mother made me do it! • The features of an organisms are due to an interaction of the individual’s genotype and environment Genetic Architecture: “sum” of the genetic effects upon a phenotype, including additive,dominance and parent-of-origin effects of several genes, pleiotropy and epistasis Different genetic architectures Different effects on the phenotype Types of Traits • Monogenic traits (rare) • Discrete binary characters • Modified by genetic and environmental background • Polygenic traits (common) • Discrete (e.g. bristle number on flies) or continuous (human height) -

Lesson Plan Mendelian Inheritance
Dolan DNA Learning Center Mendelian Inheritance __________________________________________________________________________________________ Overview • Computer with internet access This 90 minute lesson (two class periods of 45 minutes) is an introduction to Mendel’s Laws of Inheritance for students in Lesson Structure grades 5 through 8. By studying inherited traits in humans such as tasting PTC paper and inherited traits in plants such as Pre-lab (45 minutes) – Day 1 maize, we can understand how traits are passed down through Teacher Prep generations. A discussion of dominant and recessive traits in humans will encourage students to further explore their • Become familiar with Lab Center inheritance as well as their family inheritance. http://www.dnalc.org/labcenter/mendeliangenetics/m endeliangenetics_d.html Learning Outcomes • Print and copy Background Reading from the Students will be able to: Student Lab Notebook on the Lab center. • discuss the contributions of Gregor Mendel and his • Print and copy Student Pre-lab Worksheets from experiments with the garden pea. the Student Lab Notebook on the Lab center. • review the structure of DNA and chromosomes. • Cut paper strips for Sentence Strips activity. • compare a dominant trait to a recessive trait. • Make sure computers with Internet access are • compare a homozygous trait to a heterozygous trait. available. • identify traits in themselves that are either dominant or recessive. Before class • use maize as a model organism to study Mendelian Students will receive the background reading to read for inheritance. homework the night before starting lab. They will write 2 to 3 • demonstrate Mendel’s Law of Dominance and Law questions they have about the background information. -
Natural Selection One of the Most Important Contributions Made to The
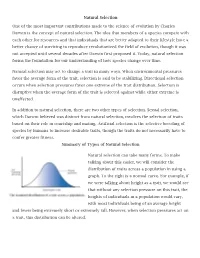
Natural Selection One of the most important contributions made to the science of evolution by Charles Darwin is the concept of natural selection. The idea that members of a species compete with each other for resources and that individuals that are better adapted to their lifestyle have a better chance of surviving to reproduce revolutionized the field of evolution, though it was not accepted until several decades after Darwin first proposed it. Today, natural selection forms the foundation for our understanding of how species change over time. Natural selection may act to change a trait in many ways. When environmental pressures favor the average form of the trait, selection is said to be stabilizing. Directional selection occurs when selection pressures favor one extreme of the trait distribution. Selection is disruptive when the average form of the trait is selected against while either extreme is unaffected. In addition to natural selection, there are two other types of selection. Sexual selection, which Darwin believed was distinct from natural selection, involves the selection of traits based on their role in courtship and mating. Artificial selection is the selective breeding of species by humans to increase desirable traits, though the traits do not necessarily have to confer greater fitness. Summary of Types of Natural Selection Natural selection can take many forms. To make talking about this easier, we will consider the distribution of traits across a population in using a graph. To the right is a normal curve. For example, if we were talking about height as a trait, we would see that without any selection pressure on this trait, the heights of individuals in a population would vary, with most individuals being of an average height and fewer being extremely short or extremely tall. -

Detecting the Form of Selection from DNA Sequence Data
Outlook COMMENT Male:female mutation ratio muscular dystrophy, neurofibromatosis and retinoblast- outcome of further investigation, this unique transposition oma), there is a large male excess. So, I believe the evidence event and perhaps others like it are sure to provide impor- is convincing that the male base-substitution rate greatly tant cytogenetic and evolutionary insights. exceeds the female rate. There are a number of reasons why the historical Acknowledgements male:female mutation ratio is likely to be less than the I have profited greatly from discussions with my colleagues current one, the most important being the lower age of B. Dove, B. Engels, C. Denniston and M. Susman. My reproduction in the earliest humans and their ape-like greatest debt is to D. Page for a continuing and very fruitful ancestors. More studies are needed. But whatever the dialog and for providing me with unpublished data. References Nature 406, 622–625 6 Engels, W.R. et al. (1994) Long-range cis preference in DNA 1 Vogel, F. and Motulsky, A. G. (1997) Human Genetics: 4 Shimmin, L.C. et al. (1993) Potential problems in estimating homology search over the length of a Drosophila chromosome. Problems and Approaches. Springer-Verlag the male-to-female mutation rate ratio from DNA sequence Science 263, 1623–1625 2 Crow, J. F. (1997) The high spontaneous mutation rate: is it a data. J. Mol. Evol. 37, 160–166 7 Crow, J.F. (2000) The origins, patterns and implications of health risk? Proc. Natl. Acad. Sci. U. S. A. 94, 8380–8386 5 Richardson, C. et al. -

The Peppered Moth and Industrial Melanism: Evolution of a Natural Selection Case Study
Heredity (2013) 110, 207–212 & 2013 Macmillan Publishers Limited All rights reserved 0018-067X/13 www.nature.com/hdy REVIEW The peppered moth and industrial melanism: evolution of a natural selection case study LM Cook1 and IJ Saccheri2 From the outset multiple causes have been suggested for changes in melanic gene frequency in the peppered moth Biston betularia and other industrial melanic moths. These have included higher intrinsic fitness of melanic forms and selective predation for camouflage. The possible existence and origin of heterozygote advantage has been debated. From the 1950s, as a result of experimental evidence, selective predation became the favoured explanation and is undoubtedly the major factor driving the frequency change. However, modelling and monitoring of declining melanic frequencies since the 1970s indicate either that migration rates are much higher than existing direct estimates suggested or else, or in addition, non-visual selection has a role. Recent molecular work on genetics has revealed that the melanic (carbonaria) allele had a single origin in Britain, and that the locus is orthologous to a major wing patterning locus in Heliconius butterflies. New methods of analysis should supply further information on the melanic system and on migration that will complete our understanding of this important example of rapid evolution. Heredity (2013) 110, 207–212; doi:10.1038/hdy.2012.92; published online 5 December 2012 Keywords: Biston betularia; carbonaria gene; mutation; predation; non-visual selection; migration INTRODUCTION EARLY EVIDENCE OF CHANGE The peppered moth Biston betularia (L.) and its melanic mutant will The peppered moth was the most diagrammatic example of the be familiar to readers of Heredity as an example of rapid evolutionary phenomenon of industrial melanism that came to be recognised in change brought about by natural selection in a changing environment, industrial and smoke-blackened parts of England in the mid-nine- evenifthedetailsofthestoryarenot.Infact,thedetailsarelesssimple teenth century.