Abstracts and Program. – 9Th International Symposium Cephalopods ‒ Present and Past in Combination with the 5Th
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Invertebrate Paleontology Devonian Goniatites from Nevada
N. Jb. Geol. Palaont. Abh. 122 3 337—342 Stuttgart. Juni 1965 Invertebrate Paleontology Earth Sciences Division Natural History Museum Devonian goniatites from Nevada By M. R. House University Museum, Oxford With plate 32 Abstract: Several goniatites recently found in the Nevada Limestone and Pilot Shale * Nevada are described. A new species, Erbenoceras erbeni, is erected for specimens from *< Lower Nevada Limestone in the Cortez Mountains and is thought to be of Emsian age. -im higher in the Nevada Limestone in the southern Roberts Mountains, but below the »3-known Siringocephahts-hezting level, occurs Cabrieroceras aff. crispiforme, which would -Tear to indicate a basal Givetian horizon. A poorly preserved lmitoceras is recorded ~tn the Pilot Shale in the Pahranagat Range. All these three genera are recorded from «Tada for the first time. Comments are made on the palaeogeographic significance of the Tjrrence of these distinctively European forms in the western United States. Introduction Through the kindness of Dr. J. G. JOHNSON of the University of Cali- nia at Los Angeles, several Devonian goniatite faunas have been sub- *rted for study and are reported on here. They include the first Middle Vvonian goniatites known from the western United States, and these serve * give a tentative placing of the Eifelian/Givetian boundary in the Nevada -nestone. Also there are two particularly interesting Lower Devonian rcimens which must be from near the level which has yielded the holotype ' Agoniatites nevadensis MILLER, which has recently been considered to be a '-cherticeras (HOUSE 1962, p. 262) and that which yielded Goniatites desidera- * WALCOTT which Dr. -

Nautiloid Shell Morphology
MEMOIR 13 Nautiloid Shell Morphology By ROUSSEAU H. FLOWER STATEBUREAUOFMINESANDMINERALRESOURCES NEWMEXICOINSTITUTEOFMININGANDTECHNOLOGY CAMPUSSTATION SOCORRO, NEWMEXICO MEMOIR 13 Nautiloid Shell Morphology By ROUSSEAU H. FLOIVER 1964 STATEBUREAUOFMINESANDMINERALRESOURCES NEWMEXICOINSTITUTEOFMININGANDTECHNOLOGY CAMPUSSTATION SOCORRO, NEWMEXICO NEW MEXICO INSTITUTE OF MINING & TECHNOLOGY E. J. Workman, President STATE BUREAU OF MINES AND MINERAL RESOURCES Alvin J. Thompson, Director THE REGENTS MEMBERS EXOFFICIO THEHONORABLEJACKM.CAMPBELL ................................ Governor of New Mexico LEONARDDELAY() ................................................... Superintendent of Public Instruction APPOINTEDMEMBERS WILLIAM G. ABBOTT ................................ ................................ ............................... Hobbs EUGENE L. COULSON, M.D ................................................................. Socorro THOMASM.CRAMER ................................ ................................ ................... Carlsbad EVA M. LARRAZOLO (Mrs. Paul F.) ................................................. Albuquerque RICHARDM.ZIMMERLY ................................ ................................ ....... Socorro Published February 1 o, 1964 For Sale by the New Mexico Bureau of Mines & Mineral Resources Campus Station, Socorro, N. Mex.—Price $2.50 Contents Page ABSTRACT ....................................................................................................................................................... 1 INTRODUCTION -

Siphuncular Structure in the Extant Spirula and in Other Coleoids (Cephalopoda)
GFF ISSN: 1103-5897 (Print) 2000-0863 (Online) Journal homepage: http://www.tandfonline.com/loi/sgff20 Siphuncular Structure in the Extant Spirula and in Other Coleoids (Cephalopoda) Harry Mutvei To cite this article: Harry Mutvei (2016): Siphuncular Structure in the Extant Spirula and in Other Coleoids (Cephalopoda), GFF, DOI: 10.1080/11035897.2016.1227364 To link to this article: http://dx.doi.org/10.1080/11035897.2016.1227364 Published online: 21 Sep 2016. Submit your article to this journal View related articles View Crossmark data Full Terms & Conditions of access and use can be found at http://www.tandfonline.com/action/journalInformation?journalCode=sgff20 Download by: [Dr Harry Mutvei] Date: 21 September 2016, At: 11:07 GFF, 2016 http://dx.doi.org/10.1080/11035897.2016.1227364 Siphuncular Structure in the Extant Spirula and in Other Coleoids (Cephalopoda) Harry Mutvei Department of Palaeobiology, Swedish Museum of Natural History, Box 50007, SE-10405 Stockholm, Sweden ABSTRACT ARTICLE HISTORY The shell wall in Spirula is composed of prismatic layers, whereas the septa consist of lamello-fibrillar nacre. Received 13 May 2016 The septal neck is holochoanitic and consists of two calcareous layers: the outer lamello-fibrillar nacreous Accepted 23 June 2016 layer that continues from the septum, and the inner pillar layer that covers the inner surface of the septal KEYWORDS neck. The pillar layer probably is a structurally modified simple prisma layer that covers the inner surface of Siphuncular structures; the septal neck in Nautilus. The pillars have a complicated crystalline structure and contain high amount of connecting rings; Spirula; chitinous substance. -
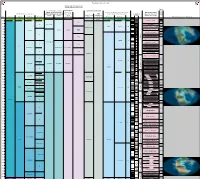
Expanded Jurassic Timescale
TimeScale Creator 2012 chart Russian and Ural regional units Russia Platform regional units Calca Jur-Cret boundary regional Russia Platform East Asian regional units reous stages - British and Boreal Stages (Jur- Australia and New Zealand regional units Marine Macrofossils Nann Standard Chronostratigraphy British regional Boreal regional Cret, Perm- Japan New Zealand Chronostratigraphy Geomagnetic (Mesozoic-Paleozoic) ofossil stages stages Carb & South China (Neogene & Polarity Tethyan Ammonoids s Ma Period Epoch Age/Stage Substage Cambrian) stages Cret) NZ Series NZ Stages Global Reconstructions (R. Blakey) Ryazanian Ryazanian Ryazanian [ no stages M17 CC2 Cretaceous Early Berriasian E Kochian Taitai Um designated ] M18 CC1 145 Berriasella jacobi M19 NJT1 Late M20 7b 146 Lt Portlandian M21 Durangites NJT1 M22 7a 147 Oteke Puaroan Op M22A Micracanthoceras microcanthum NJT1 Penglaizhenian M23 6b 148 Micracanthoceras ponti / Volgian Volgian Middle M24 Burckhardticeras peroni NJT1 Tithonian M24A 6a 149 M24B Semiformiceras fallauxi NJT15 M25 b E 150 M25A Semiformiceras semiforme NJT1 5a Early M26 Semiformiceras darwini 151 lt-Oxf N M-Sequence Hybonoticeras hybonotum 152 Ohauan Ko lt-Oxf R Kimmeridgian Hybonoticeras beckeri 153 m- Lt Late Oxf N Aulacostephanus eudoxus 154 m- NJT14 Late Aspidoceras acanthicum Oxf R Kimmeridgian Kimmeridgian Kimmeridgian Crussoliceras divisum 155 155.431 Card- N Ataxioceras hypselocyclum 156 E Early e-Oxf Sutneria platynota R Idoceras planula Suiningian 157 Cal- Oxf N Epipeltoceras bimammatum 158 lt- Lt Callo -

Download Curriculum Vitae
NEIL H. LANDMAN CURATOR, CURATOR-IN-CHARGE AND PROFESSOR DIVISION OF PALEONTOLOGY HIGHEST DEGREE EARNED Ph.D. AREA OF SPECIALIZATION Evolution, life history, and systematics of externally shelled cephalopods EDUCATIONAL EXPERIENCE Ph.D. in Geology, Yale University, 1982 M. Phil. in Geology, Yale University, 1977 M.S. in Earth Sciences, Adelphi University, 1975 B.S. in Mathematics, summa cum laude, Polytechnic University of New York, 1972 PREVIOUS EXPERIENCE IN DOCTORAL EDUCATION FACULTY APPOINTMENTS Adjunct Professor, Department of Biology, City College Adjunct Professor, Department of Geology, Brooklyn College GRADUATE ADVISEES Susan Klofak, Biology, CUNY, 1999-present Krystal Kallenberg, Marine Sciences, Stony Brook, 2003-present GRADUATE COMMITTEES Christian Soucier, Biology, Brooklyn College, 2004-present Krystal Kallenberg, Marine Sciences, Stony Brook, 2003-present Yumiko Iwasaki, Geology, CUNY, 2000-2009 Emily Allen, Geology, University of Chicago, 2002-2005 Susan Klofak, Biology, CUNY, 1996-present Claude Monnet, University of Zurich, presently Sophie Low, Geology, Harvard University RESEARCH GRANT SUPPORT Kosciuszko Foundation. Comparative study of ammonite faunas from the United States Western Interior and Polish Lowland. Post-doc: Izabela Ploch, Geological Museum of Polish Geological Institute. 2011. NSF Grant MR1-R2 (Co-PI): Acquisition of a High Resolution CT-Scanner at the American Museum of Natural History: 2010-2013. NSF Grant No. DBI 0619559 (Co-PI): Acquisition of a Variable Pressure SEM at the AMNH: 2006-2009 NSF Grant No. EAR 0308926 (PI): Collaborative Research: Paleobiology, paleoceanography, and paleoclimatology of a time slice through the Western Interior Seaway: 2003-2006 National Science Foundation, Collaborative Research: Soft Tissues and Membrane Preservation in Permian Cephalopods, $40,000, February 1, 2002-January 31, 2006. -

A Review of the Classification of Jurassic Aspidoceratid Ammonites – the Superfamily Aspidoceratoidea
VOLUMINA JURASSICA, 2020, XVIII (1): 47–52 DOI: 10.7306/VJ.18.4 A review of the classification of Jurassic aspidoceratid ammonites – the Superfamily Aspidoceratoidea Horacio PARENT1, Günter SCHWEIGERT2, Armin SCHERZINGER3 Key words: Superfamily Aspidoceratoidea, Aspidoceratidae, Epipeltoceratinae emended, Peltoceratidae, Gregoryceratinae nov. subfam. Abstract. The aspidoceratid ammonites have been traditionally included in the superfamily Perisphinctoidea. However, the basis of this is unclear for they bear unique combinations of characters unknown in typical perisphinctoids: (1) the distinct laevaptychus, (2) stout shells with high growth rate of the whorl section area, (3) prominent ornamentation with tubercles, spines and strong growth lines running in parallel over strong ribs, (4) lack of constrictions, (5) short to very short bodychamber, and (6) sexual dimorphism characterized by minia- turized microconchs and small-sized macroconchs besides the larger ones, including changes of sex during ontogeny in many cases. Considering the uniqueness of these characters we propose herein to raise the family Aspidoceratidae to the rank of a superfamily Aspi- doceratoidea, ranging from the earliest Late Callovian to the Early Berriasian Jacobi Zone. The new superfamily includes two families, Aspidoceratidae (Aspidoceratinae, Euaspidoceratinae, Epipeltoceratinae and Hybonoticeratinae), and Peltoceratidae (Peltoceratinae and Gregoryceratinae nov. subfam.). The highly differentiated features of the aspidoceratoids indicate that their life-histories -

Constraints on the Timescale of Animal Evolutionary History
Palaeontologia Electronica palaeo-electronica.org Constraints on the timescale of animal evolutionary history Michael J. Benton, Philip C.J. Donoghue, Robert J. Asher, Matt Friedman, Thomas J. Near, and Jakob Vinther ABSTRACT Dating the tree of life is a core endeavor in evolutionary biology. Rates of evolution are fundamental to nearly every evolutionary model and process. Rates need dates. There is much debate on the most appropriate and reasonable ways in which to date the tree of life, and recent work has highlighted some confusions and complexities that can be avoided. Whether phylogenetic trees are dated after they have been estab- lished, or as part of the process of tree finding, practitioners need to know which cali- brations to use. We emphasize the importance of identifying crown (not stem) fossils, levels of confidence in their attribution to the crown, current chronostratigraphic preci- sion, the primacy of the host geological formation and asymmetric confidence intervals. Here we present calibrations for 88 key nodes across the phylogeny of animals, rang- ing from the root of Metazoa to the last common ancestor of Homo sapiens. Close attention to detail is constantly required: for example, the classic bird-mammal date (base of crown Amniota) has often been given as 310-315 Ma; the 2014 international time scale indicates a minimum age of 318 Ma. Michael J. Benton. School of Earth Sciences, University of Bristol, Bristol, BS8 1RJ, U.K. [email protected] Philip C.J. Donoghue. School of Earth Sciences, University of Bristol, Bristol, BS8 1RJ, U.K. [email protected] Robert J. -

Early Ontogeny of Jurassic Bakevelliids and Their Bearing on Bivalve Evolution
Early ontogeny of Jurassic bakevelliids and their bearing on bivalve evolution NIKOLAUS MALCHUS Malchus, N. 2004. Early ontogeny of Jurassic bakevelliids and their bearing on bivalve evolution. Acta Palaeontologica Polonica 49 (1): 85–110. Larval and earliest postlarval shells of Jurassic Bakevelliidae are described for the first time and some complementary data are given concerning larval shells of oysters and pinnids. Two new larval shell characters, a posterodorsal outlet and shell septum are described. The outlet is homologous to the posterodorsal notch of oysters and posterodorsal ridge of arcoids. It probably reflects the presence of the soft anatomical character post−anal tuft, which, among Pteriomorphia, was only known from oysters. A shell septum was so far only known from Cassianellidae, Lithiotidae, and the bakevelliid Kobayashites. A review of early ontogenetic shell characters strongly suggests a basal dichotomy within the Pterio− morphia separating taxa with opisthogyrate larval shells, such as most (or all?) Praecardioida, Pinnoida, Pterioida (Bakevelliidae, Cassianellidae, all living Pterioidea), and Ostreoida from all other groups. The Pinnidae appear to be closely related to the Pterioida, and the Bakevelliidae belong to the stem line of the Cassianellidae, Lithiotidae, Pterioidea, and Ostreoidea. The latter two superfamilies comprise a well constrained clade. These interpretations are con− sistent with recent phylogenetic hypotheses based on palaeontological and genetic (18S and 28S mtDNA) data. A more detailed phylogeny is hampered by the fact that many larval shell characters are rather ancient plesiomorphies. Key words: Bivalvia, Pteriomorphia, Bakevelliidae, larval shell, ontogeny, phylogeny. Nikolaus Malchus [[email protected]], Departamento de Geologia/Unitat Paleontologia, Universitat Autòno− ma Barcelona, 08193 Bellaterra (Cerdanyola del Vallès), Spain. -

Size Distribution of the Late Devonian Ammonoid Prolobites: Indication for Possible Mass Spawning Events
Swiss J Geosci (2010) 103:475–494 DOI 10.1007/s00015-010-0036-y Size distribution of the Late Devonian ammonoid Prolobites: indication for possible mass spawning events Sonny Alexander Walton • Dieter Korn • Christian Klug Received: 17 December 2009 / Accepted: 18 October 2010 / Published online: 15 December 2010 Ó Swiss Geological Society 2010 Abstract Worldwide, the ammonoid genus Prolobites is Abbreviations only known from a few localities, and from these fossil SMF Senckenberg Museum, Frankfurt a. M., Germany beds almost all of the specimens are adults as shown by the MB.C. Cephalopod collection of the Museum fu¨r presence of a terminal growth stage. This is in marked Naturkunde Berlin (the collections of Franz contrast to the co-occurring ammonoid genera such as Ademmer, Werner Bottke, and Harald Simon Sporadoceras, Prionoceras, and Platyclymenia. Size dis- are incorporated here) tribution of specimens of Prolobites from three studied localities show that, unlike in the co-occurring ammonoid species, most of the material belongs to adult individuals. The morphometric analysis of Prolobites delphinus Introduction (SANDBERGER &SANDBERGER 1851) demonstrates the intra- specific variability including variants with elliptical coiling The mid-Fammenian (Late Devonian) ammonoid genus and that dimorphism is not detectable. The Prolobites Prolobites is only known from a few localities worldwide, material shows close resemblance to spawning populations mainly in the Rhenish Mountains and the South Urals. of Recent coleoids such as the squid Todarodes filippovae Where it does occur, the majority of the specimens show a ADAM 1975. Possible mass spawning events are discussed terminal growth stage suggesting that these are adults. -

Contributions in BIOLOGY and GEOLOGY
MILWAUKEE PUBLIC MUSEUM Contributions In BIOLOGY and GEOLOGY Number 51 November 29, 1982 A Compendium of Fossil Marine Families J. John Sepkoski, Jr. MILWAUKEE PUBLIC MUSEUM Contributions in BIOLOGY and GEOLOGY Number 51 November 29, 1982 A COMPENDIUM OF FOSSIL MARINE FAMILIES J. JOHN SEPKOSKI, JR. Department of the Geophysical Sciences University of Chicago REVIEWERS FOR THIS PUBLICATION: Robert Gernant, University of Wisconsin-Milwaukee David M. Raup, Field Museum of Natural History Frederick R. Schram, San Diego Natural History Museum Peter M. Sheehan, Milwaukee Public Museum ISBN 0-893260-081-9 Milwaukee Public Museum Press Published by the Order of the Board of Trustees CONTENTS Abstract ---- ---------- -- - ----------------------- 2 Introduction -- --- -- ------ - - - ------- - ----------- - - - 2 Compendium ----------------------------- -- ------ 6 Protozoa ----- - ------- - - - -- -- - -------- - ------ - 6 Porifera------------- --- ---------------------- 9 Archaeocyatha -- - ------ - ------ - - -- ---------- - - - - 14 Coelenterata -- - -- --- -- - - -- - - - - -- - -- - -- - - -- -- - -- 17 Platyhelminthes - - -- - - - -- - - -- - -- - -- - -- -- --- - - - - - - 24 Rhynchocoela - ---- - - - - ---- --- ---- - - ----------- - 24 Priapulida ------ ---- - - - - -- - - -- - ------ - -- ------ 24 Nematoda - -- - --- --- -- - -- --- - -- --- ---- -- - - -- -- 24 Mollusca ------------- --- --------------- ------ 24 Sipunculida ---------- --- ------------ ---- -- --- - 46 Echiurida ------ - --- - - - - - --- --- - -- --- - -- - - --- -

Newsletter 35/2, December 2008
INTERNATIONAL SUBCOMMISSION ON JURASSIC STRATIGRAPHY http://jurassic.earth.ox.ac.uk/ Newsletter 35/2, December 2008 Edited by Nicol Morton and Stephen Hesselbo Prospective GSSP for base Toarcian, Peniche, Portugal (photo: S. P. Hesselbo) CONTENTS EX-CHAIRMAN’S REPORT Nicol Morton 1 NEWS ITEMS AND MEETINGS 8th International Congress on the Jurassic System, Shehong of Suining, China August 2010 Jingeng Sha and Yongdong Wang 4 IGCP506: Marine and Non-marine Jurassic: Report on 5th International Symposium, Hammamet, Tunisia, 28-31 March 2008-11-15 Nicol Morton 5 REPORTS OF WORKING GROUPS Pliensbachian Christian Meister 7 Bathonian Sixto R. Fernandez Lopez 9 Callovian John Callomon 12 Kimmeridgian Andrzej Wierzbowski 14 Tithonian Federico Oloriz and Günther Schweigert 20 Geoconservation Kevin Page 21 Liaison Robert Chandler 27 CORRESPONDENCE Session on the Jurassic System at the 2007 annual meeting of the Geological Society of Japan. Atsushi Matsuoka 30 Annual meeting of the German Subcommission on Jurassic Stratigraphy, Bamberg (Bavaria) 22-24 May 2008. Eckhard Mönnig 31 Trans-border (S.E. Serbia/S.W. Bulgaria) correlation of Jurassic sediments – a new bilateral project. Platon Tchoumatchenko 32 BOOK ANNOUNCEMENTS Stratigraphy: Terminology and Practice. J. Rey and S. Galeotti (eds.) 2008, Technip Édit. BRGM-Total 176p. Jacques Thierry 41 Proceedings of the Geologists’ Association vol. 119, Part 1, 2008, Special Issue on the Jurassic edited by Beris M. Cox and Michael G. Sumbler Nicol Morton 42 IN MEMORIAM Larbi MEKHALI (1952-2008) Abbas Marok 43 René MOUTERDE (1915–2007) Christiane Ruget 45 Colin F. PARSONS (1945-2008) Robert Chandler 47 Bruce W. SELLWOOD (1946-2007) Tony Hallam 50 EX-CHAIRMAN’S REPORT Nicol MORTON The Jurassic Subcommission and the [email protected] international Jurassic community in general have the advantages of two changes of I am happy to report to you all that this is my personnel at the same time as the advent of last such Report in the ISJS Newsletter. -

Remarks on the Tithonian–Berriasian Ammonite Biostratigraphy of West Central Argentina
Volumina Jurassica, 2015, Xiii (2): 23–52 DOI: 10.5604/17313708 .1185692 Remarks on the Tithonian–Berriasian ammonite biostratigraphy of west central Argentina Alberto C. RICCARDI 1 Key words: Tithonian–Berriasian, ammonites, west central Argentina, calpionellids, nannofossils, radiolarians, geochronology. Abstract. Status and correlation of Andean ammonite biozones are reviewed. Available calpionellid, nannofossil, and radiolarian data, as well as radioisotopic ages, are also considered, especially when directly related to ammonite zones. There is no attempt to deal with the definition of the Jurassic–Cretaceous limit. Correlation of the V. mendozanum Zone with the Semiforme Zone is ratified, but it is open to question if its lower part should be correlated with the upper part of the Darwini Zone. The Pseudolissoceras zitteli Zone is characterized by an assemblage also recorded from Mexico, Cuba and the Betic Ranges of Spain, indicative of the Semiforme–Fallauxi standard zones. The Aulacosphinctes proximus Zone, which is correlated with the Ponti Standard Zone, appears to be closely related to the overlying Wind hauseniceras internispinosum Zone, although its biostratigraphic status needs to be reconsidered. On the basis of ammonites, radiolarians and calpionellids the Windhauseniceras internispinosum Assemblage Zone is approximately equivalent to the Suarites bituberculatum Zone of Mexico, the Paralytohoplites caribbeanus Zone of Cuba and the Simplisphinctes/Microcanthum Zone of the Standard Zonation. The C. alternans Zone could be correlated with the uppermost Microcanthum and “Durangites” zones, although in west central Argentina it could be mostly restricted to levels equivalent to the “Durangites Zone”. The Substeueroceras koeneni Zone ranges into the Occitanica Zone, Subalpina and Privasensis subzones, the A.